
アストロサイト同定のためのマーカー選択
Haddie DeHart著(Indiana University School of Medicine、博士課程在籍)
|
目次 はじめに |
はじめに
アストロサイト(星状膠細胞)は脳や脊髄の機能に極めて重要な役割を果たす、中枢神経系(CNS:Central nervous system)に存在する星状のグリア細胞です。従来、アストロサイトは単に神経細胞を補助するだけの存在と考えられていましたが、血液脳関門の制御、イオン恒常性の維持、神経伝達物質のクリアランス等においても極めて重要な役割を果たすことが明らかになっています。アストロサイトをはじめとするグリア細胞は脳を構成する細胞の約50%を占め、アストロサイトの正常な機能や神経疾患への関与は脳研究における最前線の研究テーマとなっています。それに伴い、アストロサイトそのものの同定や個々の細胞の生物学的な役割を詳細に解析可能なアストロサイト特異的なマーカーの重要性が浮き彫りになっています。
代表的なアストロサイトマーカー
アストロサイトを識別・標識するために、ウェスタンブロット(WB:Western blot)、免疫組織化学染色(IHC:Immunohistochemistry)、免疫蛍光染色(IF:Immunofluorescence)等のイムノアッセイでは、アストロサイトに特徴的なタンパク質を認識する抗体が広く用いられています。
主要なアストロサイトマーカーは以下の通りです。
GFAP(Glial Fibrillary Acidic Protein)
GFAPは細胞骨格の支持に極めて重要な役割を果たす中間径フィラメントタンパク質として広く認知されています。主にアストロサイトに発現しているため、成熟アストロサイトの代表的なマーカーとされています。さらに、中枢神経系の損傷、炎症、神経変性疾患に伴い発現量が増加するため、GFAP発現は反応性アストロサイトを同定する際の指標としても極めて有用です。しかし、特定の病態においては他の細胞でもGFAP発現が上昇する場合があるため注意を要します。したがって、アストロサイトとその他の細胞を識別するために、複数のマーカーの併用、または形態学的解析の併用が必要とされる場合があります。
プロテインテックのGFAP抗体は、標識抗体・未標識抗体ともに多彩なラインアップの中から実験系に適した抗体をご選択いただけます。
|
図1. GFAP抗体(カタログ番号:81063-1-RR、希釈倍率1:200)、CoraLite® 488-conjugated Goat Anti-Rabbit IgG (H+L)抗体を使用したラット脳組織(4% PFA固定)の免疫蛍光染色解析像。 |
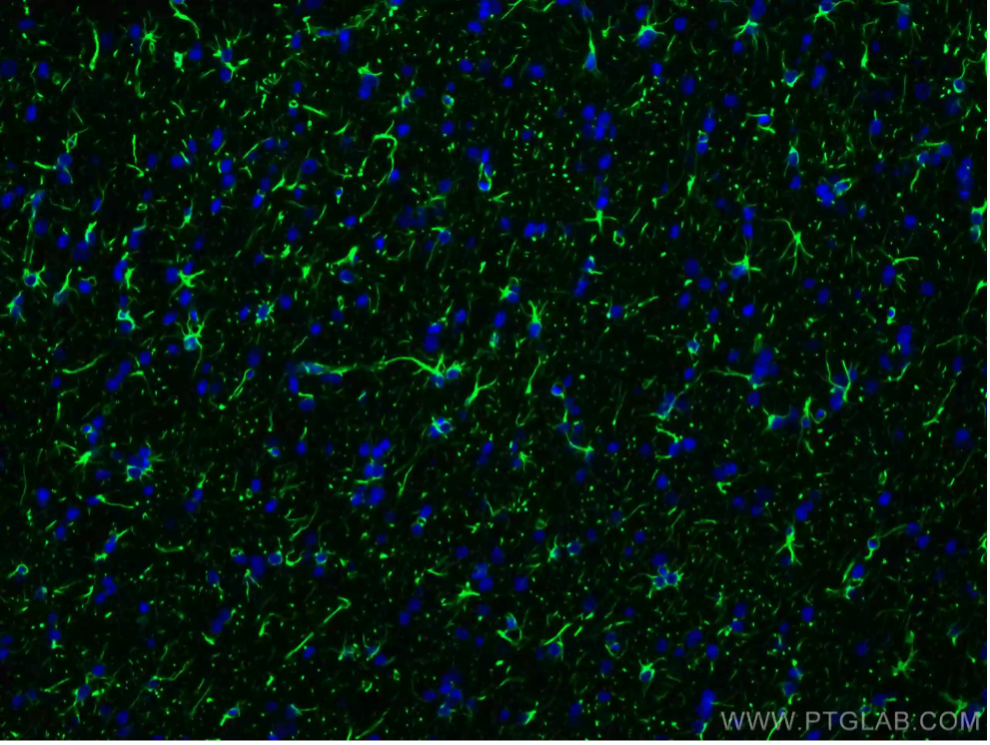
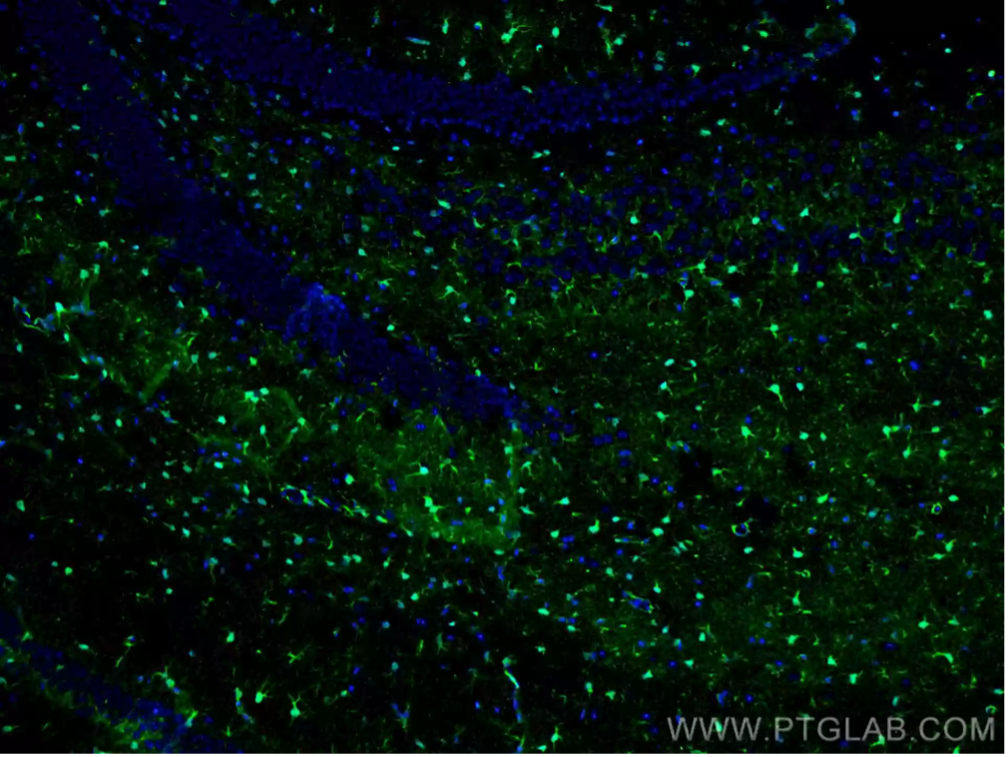
S100β(S100 Calcium-Binding Protein B)
S100βは、Ca2+シグナル伝達系、神経保護、傷害応答に極めて重要な役割を果たすカルシウム結合タンパク質です。主に未成熟アストロサイトや成熟アストロサイトに発現し、脳組織の状態を研究するにあたり有用な情報を得られるマーカーとなります。また、正常な組織や疾患等の特定の条件下にあるアストロサイトの反応性を評価するためにS100βの発現レベルを解析することがあります。
プロテインテックはS100β検出用抗体をはじめ、必要な抗体・試薬類がすべて揃ったIHCキット「IHCeasy」や各種ELISAキットを取り揃えています。
|
図2. S100B抗体(カタログ番号:15146-1-AP、希釈倍率1:200)、CoraLite® 488-conjugated Goat Anti-Rabbit IgG (H+L)抗体を使用したマウス脳組織(4% PFA固定)の免疫蛍光染色解析像。 |
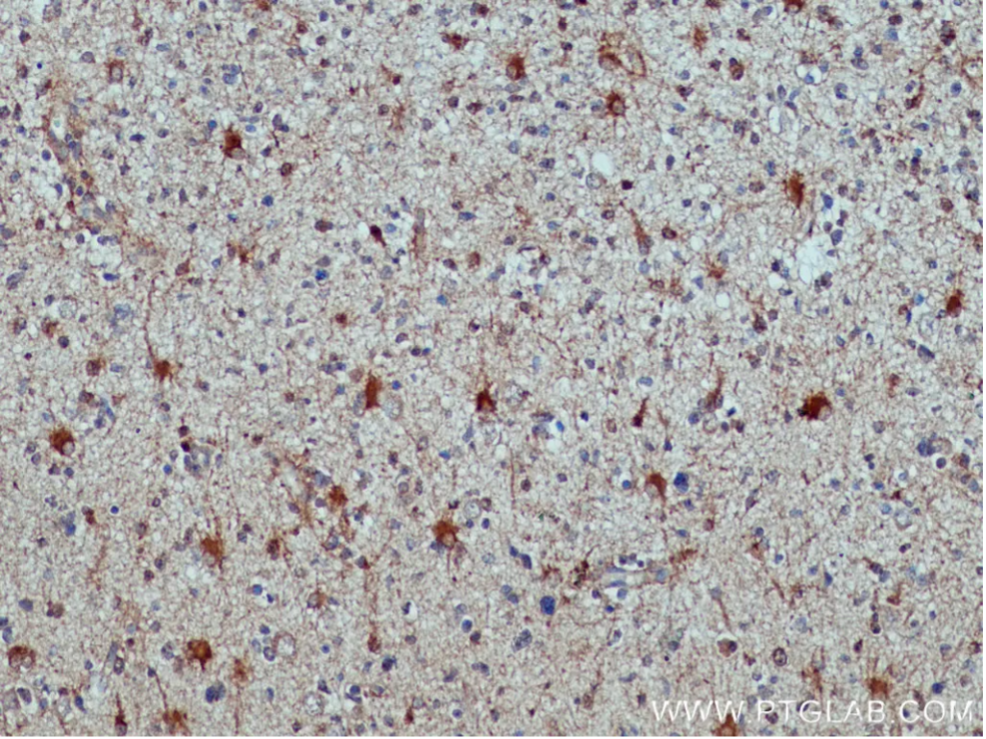
ALDH1L1(Aldehyde Dehydrogenase 1 Family Member L1)
アストロサイトの遺伝子発現プロファイルの解析によって、ALDH1L1をはじめとする幅広いアストロサイトマーカーの同定が進んでいます。ALDH1L1は、細胞代謝の中でも特にアルデヒドの解毒や葉酸代謝において極めて重要な役割を果たします。成熟脳の汎アストロサイトマーカーとして知られ、アストロサイトに極めて特異的に発現するため、複数のグリア細胞集団が混在する実験系において理想的なマーカーとして機能します。
プロテインテックは、独自の厳しい検証基準をクリアした高品質なALDH1L1モノクローナル抗体・ポリクローナル抗体を取り揃えています。
|
図3. ALDH1L1抗体(カタログ番号:17390-1-AP、希釈倍率1:200)を使用したパラフィン包埋ヒト神経膠腫スライドの免疫組織化学染色。Tris-EDTA buffer(pH 9.0)で熱処理し抗原賦活化した試料を使用。 |
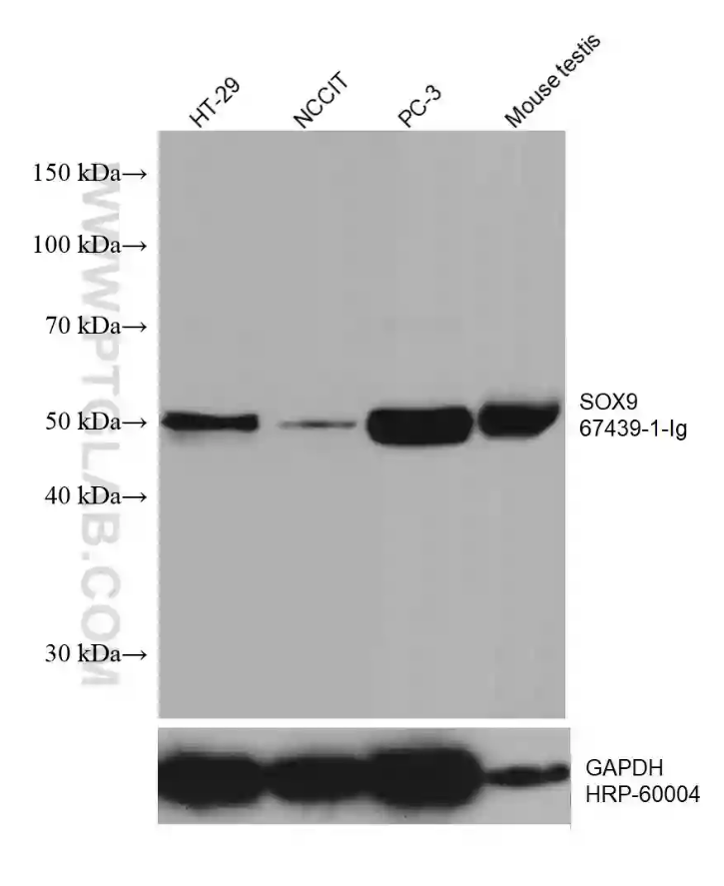
SOX9(SRY-box transcription factor 9)
神経の発生過程において、SOX9は神経前駆細胞のアストロサイトへの分化を誘導することから、アストロサイト系統の細胞の確立や維持における中心的な分子として位置づけられています。成熟アストロサイトでは、SOX9はグリア細胞の特性に結びつく遺伝子を制御し、恒常性の維持や神経の支持を促す細胞機能に貢献しています。神経細胞やその他のグリア細胞と比較して、SOX9は主にアストロサイトに高発現するため、発生過程や病態におけるアストロサイト細胞群の評価、その分化や反応性を解析する研究において信頼性の高いマーカーとなります。
プロテインテックはウェスタンブロット・IHC・フローサイトメトリーで検証済みの高感度なSOX9抗体を取り扱っています。
|
図4. SDS-PAGEに供した後、SOX9抗体(カタログ番号:67439-1-Ig、希釈倍率1:10000、室温で1.5時間インキュベーション)を使用した各種ライセートのウェスタンブロット。SOX9を検出後、メンブレンをストリッピング(抗体除去)し、ローディングコントロールとしてHRP-conjugated GAPDH モノクローナル抗体でリプローブした。 |
その他のマーカー
研究目的・研究背景に基づき、アストロサイトの機能や形態の解析に各種マーカーが用いられています。
-
-
EAAT2(Excitatory Amino Acid Transporter 2、別名:GLT-1)
-
AQP4(Aquaporin 4)
-
Cx43(Connexin 43)
-
NF-κB(Nuclear Factor Kappa-light-chain-enhancer of Activated B cells)
-
IL-6(Interleukin 6)
-
IL-1β(Interleukin 1 Beta)
-
TNF-α(Tumor Necrosis Factor Alpha)
-
CCL2(C-C Motif Chemokine Ligand 2、別名:MCP-1)
-
CXCL10(C-X-C Motif Chemokine Ligand 10、別名:IP-10)
-
CXCL12(C-X-C Motif Chemokine Ligand 12、別名:SDF-1)
-
|
病態 |
アストロサイトの挙動 |
|
アルツハイマー病(AD) |
· アミロイド-β(Aβ)やタウの病変がEAAT2の発現を抑制し、グルタミン酸の調節不全やシナプス毒性の原因となります。 · 血管周囲におけるAQP4局在の極性が喪失すると脳脊髄液(CSF)流動が障害され、アルツハイマー病発症につながります。 · Aβの沈着によりCx43の発現上昇が誘導され、アストロサイトのカリウム緩衝作用やグルタミン酸緩衝作用が損なわれ、シナプスの過活動や神経変性が生じます。 · アルツハイマー病やパーキンソン病において、反応性アストロサイトのNF-κBは過剰に活性化され、炎症誘発性サイトカイン(IL-1β、TNF-α)が産生され、神経細胞傷害を悪化させます。 |
|
筋萎縮性側索硬化症(ALS) |
· EAAT2の発現が選択的に低下することでグルタミン酸が蓄積し、運動ニューロンが死滅します。 |
|
悪性脳腫瘍 · 膠芽腫(GBM) · 高悪性度神経膠腫 · 星細胞腫 · 髄芽腫(MB) |
· AQP4の発現が亢進すると細胞内外への水の流動が活発になり、腫瘍細胞は微小環境に急速に適応し、周辺組織へ浸潤します。 · Cx43は悪性脳腫瘍において高発現し、腫瘍増殖と浸潤を促進します。 · CCL2やCXCL12は腫瘍関連マクロファージ(TAM)やミクログリアを動員し、免疫抑制性の腫瘍微小環境(TME)形成を促進します。また、IL-6やTNF-αはグリオーマ幹細胞を維持し、放射線抵抗性を高めます。 · ビメンチンはGFAPと共に腫瘍関連アストロサイトにおいて発現が亢進し、反応性アストロサイトのマーカーとして利用されます。 · 腫瘍細胞はアストロサイトのNF-κBを活性化し、IL-6、IL-1β、TNF-α等の腫瘍形成を促進するサイトカイン類の分泌を誘導します。 · アストロサイトにおけるNF-κBの活性化はがん細胞の増殖を促進し、治療抵抗性を高めます。 |
|
脳梗塞(虚血性脳卒中) |
· EAAT1/EAAT2機能不全によってトランスポーター機能が低下しグルタミン酸が蓄積すると、その興奮毒性によって神経細胞が死滅します。 · AQP4の発現亢進に伴いアストロサイトが膨張し頭蓋内圧が上昇することで、神経細胞の損傷が引き起こされます。 · 低酸素状態に陥りCx43の脱リン酸化が進むことで、ヘミチャネルの開口、細胞膨張、炎症反応が促進されます。 |
|
多発性硬化症(MS) |
· MSモデルの研究において、Cx43機能の阻害によって神経炎症が低減し再髄鞘化が促進されることが明らかになっています。 · NF-κBの活性化はCXCL10やCCL2の発現を亢進し、中枢神経系へのT細胞浸潤を促進します。 |
|
視神経脊髄炎スペクトラム障害(NMOSD) |
· 自己抗体であるNMO-IgG(抗AQP4抗体)とAQP4の結合によりアストロサイト傷害、炎症細胞浸潤、脱髄が引き起こされます。 |
|
統合失調症 |
· EAAT1/EAAT2機能不全とグルタミン酸恒常性の異常との関連が報告されています。 |
|
脳卒中 |
· EAAT1/EAAT2の機能不全により興奮性神経伝達の持続時間が延長し、発作感受性が増大します。 · AQP4はアストロサイトのグルタミン酸トランスポーターであるEAAT2と相互作用します。AQP4機能不全はグルタミン酸の取込みを抑制し、過剰興奮が引き起こされます。 · Cx43ヘミチャネルの活性化により神経炎症・興奮毒性・二次的な神経組織損傷が生じ、てんかん発症を促進します。また、Cx43の特定の部位のリン酸化・脱リン酸化の変化が外傷後のてんかん発症に関連しています。 · 脳の傷害後、反応性アストロサイトにおけるビメンチン発現が亢進します。この発現亢進はアストロサイトの肥大とグリア瘢痕形成を促します。 · IL-1βやTNF-αの過剰分泌は、アストロサイトのグルタミン酸取込み機構を障害し、興奮毒性を悪化させます。 · IL-6、TNF-α、CXCL10等の分泌サイトカインがタイトジャンクションを脆弱化し、血液脳関門(BBB)の透過性を亢進させます。 |
マーカー選択時に考慮すべきポイント
どのような研究であれ、研究の背景を理解し、研究に適したマーカーを選択することが重要です。その際、留意すべき主なポイントは以下の通りです。
- 研究対象のアストロサイトの脳内における領域特異性
- アストロサイトは分子生物学的研究や機能解析により、脳全体でみると不均一であることが示されています。
- マウスの場合、GFAPは海馬と中脳のアストロサイトでより多く発現しているのに対し、大脳や後脳のアストロサイトでは相対的にS100βの優勢な発現が認められます。
- どのような状態にあるアストロサイトに注目するか
- GFAPは一般的に成熟・分化が進んだアストロサイトに豊富に存在する傾向にあります。アストロサイトの発達過程を研究する場合は、S100βに加え初期段階から発現が認められるEAAT1に注目するという選択肢も検討しましょう。
- 使用する動物種
- ヒト・マウス・ラット等のモデル動物のアストロサイトにおける種特有の違いに注意してください。
- Liらによる論文(Li, et al. 2021)では、ヒトおよびマウスのアストロサイトのトランスクリプトームの比較解析が行われています。遺伝子発現プロファイルの大部分は種間で保存されていましたが、マウスアストロサイトでは代謝やミトコンドリアに関連する遺伝子が高く発現していたのに対し、ヒトでは防御反応や細胞外移行タンパク質に関連する遺伝子が高く発現していました。
- その他の細胞におけるマーカー発現
- S100βはアストロサイトマーカーとして利用頻度の高い分子ですが、オリゴデンドロサイトやオリゴデンドロサイト前駆細胞を同定するマーカーでもあります。同様に、ある種のニューロンではEAAT2が検出されます。
- 星細胞腫は悪性度が高くなるにつれALDH1L1・EAAT2・Cx43等のアストロサイトマーカーの発現が低下することが示されています。そのため、膠芽腫(GBM)におけるアストロサイトを研究する場合はこれらのマーカーの消失を1つの指標として使用することができます。また、グリオーマ組織ではGAPDHの発現量が変動しやすく、正常組織とは異なる挙動を示すためコントロールには推奨されません。
- アストロサイト同定アッセイの選択基準と留意点
- アストロサイトの形態学的解析の研究に幅広く利用されているのは、免疫蛍光染色ですが、研究のアプローチや利用可能な設備、予算に応じて別の手法も用いられます。
- 例えば、ウェスタンブロット、免疫蛍光染色、免疫組織化学の各アプリケーションで検証済みの抗体を用意すれば、1種類の抗体だけでGFAPタンパク質の発現を確認したり、培養細胞やオルガノイドにおけるアストロサイトの形態を解析したり、患者の腫瘍組織に存在するアストロサイト数を計測できます。
参考文献
- Z Yang, K K W Wang. Glial fibrillary acidic protein: from intermediate filament assembly and gliosis to neurobiomarker. Trends Neurosci. 2015 Jun;38(6):364-74.
- J Schiweck, B J Eickholt, K Murk. Important Shapeshifter: Mechanisms Allowing Astrocytes to Respond to the Changing Nervous System During Development, Injury and Disease. Front Cell Neurosci. 2018 Aug 21:12:261.
- H Zhang, et al. Novel insights into astrocyte-mediated signaling of proliferation, invasion and tumor immune microenvironment in glioblastoma. Biomed Pharmacother. 2020 Jun:126:110086.
- F Endo, et al. Molecular basis of astrocyte diversity and morphology across the CNS in health and disease. Science. 2022 Nov 4;378(6619):eadc9020.
- K A Guttenplan, S A Liddelow. Astrocytes and microglia: Models and tools. J Exp Med. 2019 Jan 7;216(1):71-83.
- M Brandao, T Simon, G Critchley, G Giamas. Astrocytes, the rising stars of the glioblastoma microenvironment. Glia. 2019 May;67(5):779-790.
- A Mega, et al. Astrocytes enhance glioblastoma growth. Glia. 2020 Feb;68(2):316-327.
- S A Liddelow, et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature. 2017 Jan 26;541(7638):481-487.
- X Yu, J Nagai, et al. Improved tools to study astrocytes. Nat Rev Neurosci. 2020 Mar;21(3):121-138.
- Y Zhang, K Chen, S A Sloan, B A Barres, J Q Wu,et al. An RNA-sequencing transcriptome and splicing database of glia, neurons, and vascular cells of the cerebral cortex. J Neurosci. 2014 Sep 3;34(36):11929-47.
- N A Oberheim, et al. Uniquely hominid features of adult human astrocytes. J Neurosci. 2009 Mar 11;29(10):3276-87.
- J Li, et al. Conservation and divergence of vulnerability and responses to stressors between human and mouse astrocytes. Nat Commun. 2021 Jun 25;12(1):3958.