
RNAのm5C修飾とがん
エピジェネティックなm5C修飾が制御するRNAの細胞内動態やがんの予後との関係について解説します。
5-メチルシトシン(m5C)修飾はエピジェネティクスの分野における化学修飾の一種で、核酸塩基であるシトシン環の5位の炭素にメチル基が付加します。この修飾は動的かつ可逆的で、生体内の多様なRNAに広範に分布し、様々なRNA分子のプロセシング・核外輸送・安定性等の動態に影響を及ぼします。
RNAに対するm5C修飾の効果
tRNA
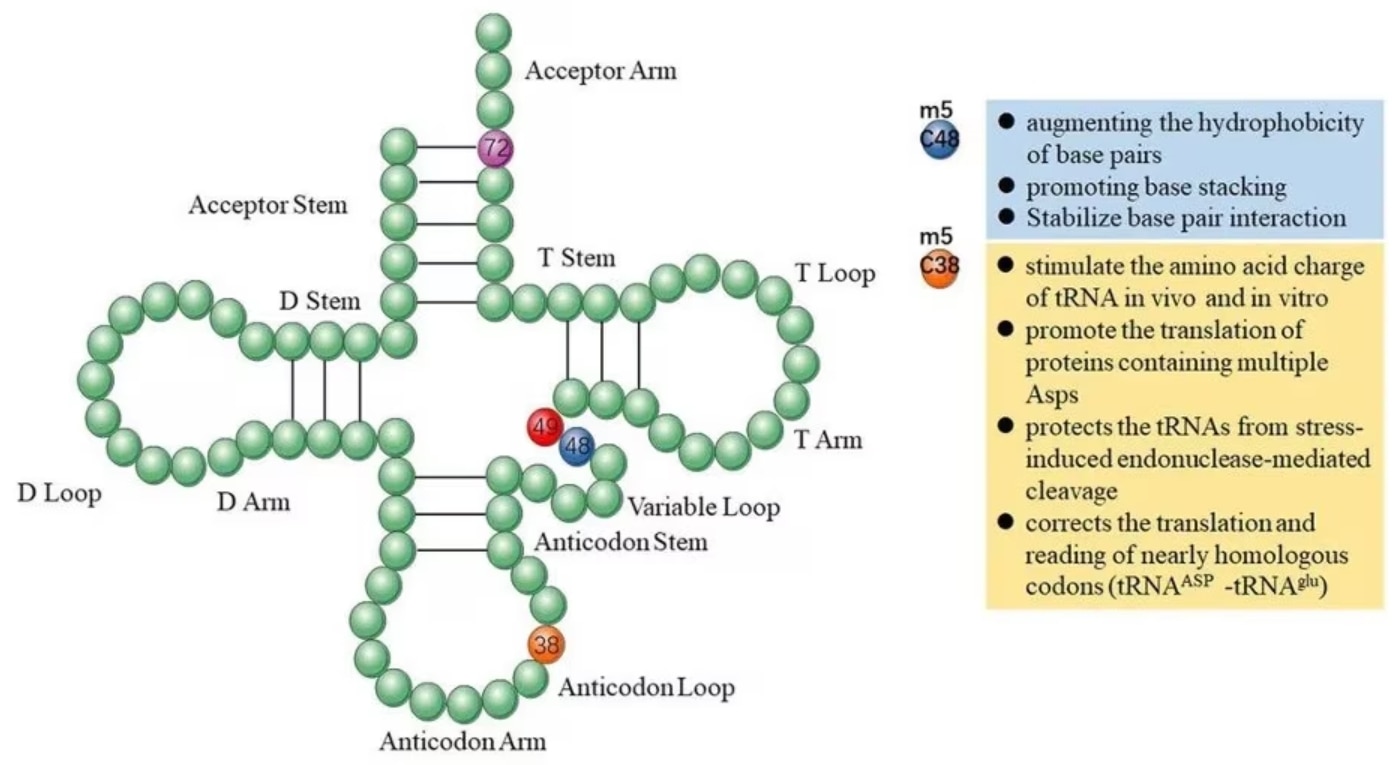
tRNA(転移RNA)の場合、m5C修飾は正確なコドン認識や安定的な塩基対形成を促し、効率的かつ正確性の高い翻訳制御、tRNAの安定性制御、細胞代謝やストレス応答に関与します[2]。マウスtRNAAspに存在するアンチコドンループのC38のメチル化はAsp-AspのようなAspが連続する配列のタンパク質の翻訳を促進するとともに、ストレス誘発性のエンドヌクレアーゼによる切断からtRNAを保護します[3]。C48のメチル化は塩基の疎水性を高め、塩基のスタッキングを促してtRNAの三次構造の安定化に寄与し、タンパク質合成効率を高めます[4]。
|
図1. tRNAのm5C修飾が果たす機能(引用元:Song et al. 2022[2]) |
rRNA
rRNA(リボソームRNA)の場合は、m5C修飾によりrRNAのコンホメーションが安定化します。酵母では酸化ストレス下にあるとき、rRNAのm5C修飾はrRNAのフォールディングを促し、様々なシグナル伝達経路に関与するmRNAの選択的リクルートメントと翻訳を促進します[5]。
lncRNA
lncRNA(長鎖ノンコーディングRNA)の遺伝子領域におけるメチル化修飾分布やその制御に関する研究も進められており、hm5C(5-ヒドロキシメチルシトシン)修飾はlncRNA転写活性化と正の相関があることが判明しています。また、ある種のがん細胞においてはhm5CがRNA Pol IIを介した長距離のクロマチン相互作用に関与してlncRNA転写が制御されます。さらに、様々なエピジェネティック修飾制御因子によるm5C修飾やhm5C修飾と、それに伴うlncRNAの転写制御が腫瘍の発生と関連しています[6]。
mRNA
以前は、mRNA(メッセンジャーRNA)はtRNAやrRNAと比較して相対的に存在量が少ないことに加え、効果的な単離・精製技術が確立されておらず、その研究は比較的進んでいませんでした。しかし、近年の検出手法の進歩に伴い、mRNAのm5C修飾に関する研究は徐々に増加しています。
mRNAのm5C修飾には、NSUN2(NOP2/SUN RNA methyltransferase 2)、NSUN6、TRDMT1(tRNA aspartate methyltransferase 1)、ALYREF(ALY/REF export factor)等の、様々な酵素やエフェクター分子が関与します[1, 7]。mRNAがm5C修飾をうけることで、mRNAの核‐細胞質間輸送の促進、ウイルスタンパク質の発現、DNA損傷修復、mRNAの安定化、増殖・遊走、幹細胞等の発生、mRNAのスプライシング制御といった多様なプロセスが制御されます。
細胞の種類やmRNAの修飾部位の違いによって異なるm5C修飾の分布は、固有の調節機能を発揮し、翻訳の促進または阻害作用を示します[1]。したがって、mRNAの異常なm5C修飾は、がんや自己免疫疾患を含む多様な疾患の発症と深く関わっています[8]。
m5C修飾の制御因子
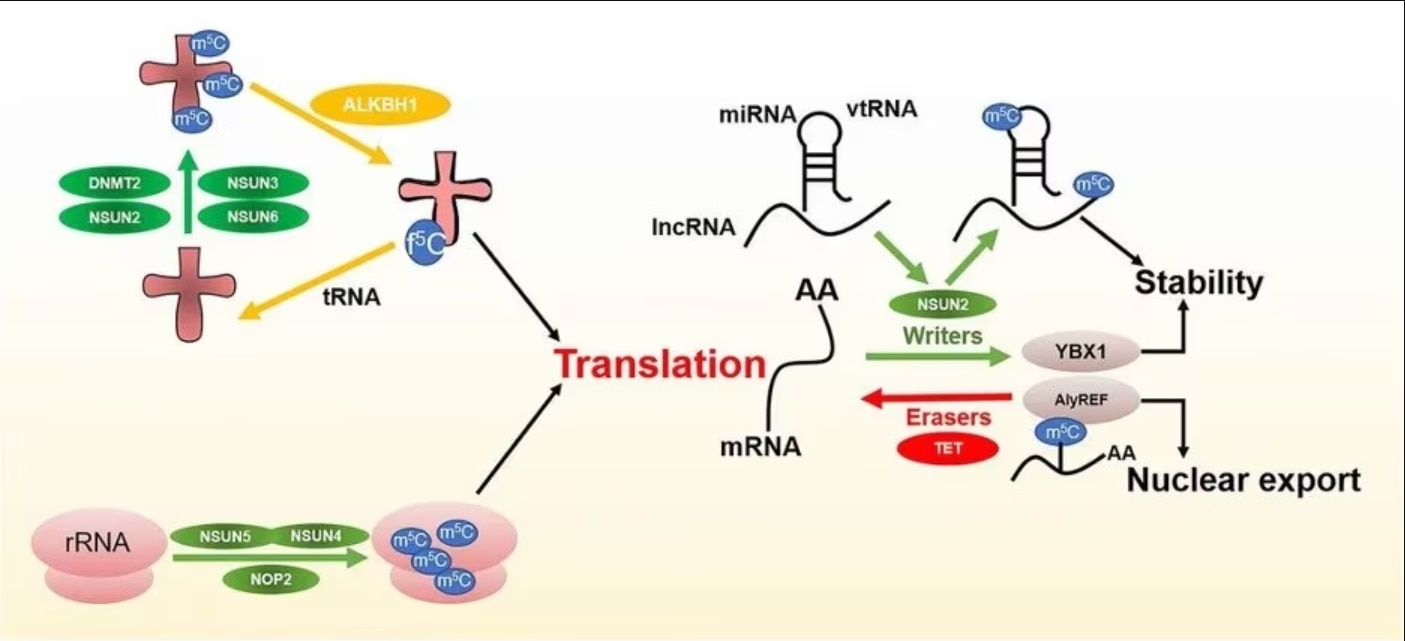
RNAのメチル化には、3種類の制御因子が主に関与します。1)WriterはRNAに特定の化学基を付加する役割を担い、2)ReaderはRNA修飾部位に結合してRNAの安定性維持と翻訳・スプライシングに関与し、3)EraserはRNA修飾を除去します。
|
図2. m5C制御因子とその機能(画像引用元:Song et al. 2022[2]) |
m5C writer
m5C writerであるメチルトランスフェラーゼは、S-アデノシルメチオニンをメチル基の供給源として用い、シトシンのピリミジン環へのメチル基転移を触媒します。
核酸のm5C修飾を担うメチルトランスフェラーゼとして、DNMT1(DNA methyltransferase 1)、DNMT1との相同性から当初はDNMT2と称されていたものの後にtRNAメチル化酵素であることが判明したTRDMT1、NSUN(NOL1/NOP2/SUN domain)タンパク質ファミリーといった数種類の酵素が明らかにされています。
NSUN2のターゲットは比較的広範にわたり、mRNA・miRNA・tRNAのメチル化で主要な役割を果たします[9]。TRDMT1はtRNAのm5C修飾を触媒しますが、異なる複数の部位のシトシン残基をメチル化するNSUN2とは異なりtRNAのアンチコドンループに存在する単一のシトシン残基を特異的にメチル化します[12]。
m5C reader
m5C修飾に応じた様々な生体反応が進行するには、修飾部位を特異的に認識・結合するreaderタンパク質の関与が欠かせません。とはいえ、m5C readerに関する研究は現在も初期段階に留まっています。
ALYREFは、核内でmRNAを認識する(読み取る)タンパク質で、mRNAを核質から核外に輸送する役割を果たします[7]。YBX1も、細胞質においてmRNAのm5C修飾を認識するタンパク質として同定され、m5C修飾mRNAの安定性を維持する役割を果たします[1]。さらに、RAD52はm5C修飾RNA-DNAハイブリッド鎖に高い親和性を示すことが認められており、RAD52がDNA損傷部位のm5C修飾を認識する働きがあることを示しています[10]。
m5C eraser
メチル化等の核酸修飾には可逆的なプロセスが存在し、多くの修飾が動的な制御をうけます。しかし、脱メチル化酵素(eraser)の分類や定義は十分に解明されていません。
DNAのm5C修飾に関連する脱メチル化プロセスは、これまでの初期研究によって既に詳細が特定されています。TETファミリーはジオキシゲナーゼとして作用し、m5Cを段階的に酸化して5caC(5-カルボキシシトシン)に変換します。5caCはチミンDNAグリコシラーゼの塩基除去作用によって切り出され、欠落した部位がシトシンに置換されることでDNAの脱メチル化が完了します。
α-ケトグルタル酸・鉄イオン依存的にTETファミリーが触媒するm5Cからhm5Cへの変換と類似のプロセスがRNAに対しても行われることが、Fuらの研究で明らかになっています。hm5Cは、TETファミリーの作用で5fC(5-ホルミルシトシン)および5caC(5-カルボキシシトシン)に変換され、最終的に塩基除去修復経路のような作用でシトシンに変換され脱メチル化される可能性が推測されています[11]。
その他に、ALKBH1という別のDNA/RNAジオキシゲナーゼがtRNAの脱メチル化に関与することが判明しています。ALKBH1の欠失によりミトコンドリア翻訳や酸素消費の重篤な障害が生じることから、m5C修飾がミトコンドリアの活動を制御している可能性が示唆されています[13, 14]。
m5C修飾とがん
近年の研究により、RNAのm5C修飾は様々な疾患の病態に関連し、特に腫瘍形成における重要性が強く示唆されています。
|
図3. がんにおけるm5C修飾制御因子の異常(画像引用元:Song et al. 2022[2])。
|
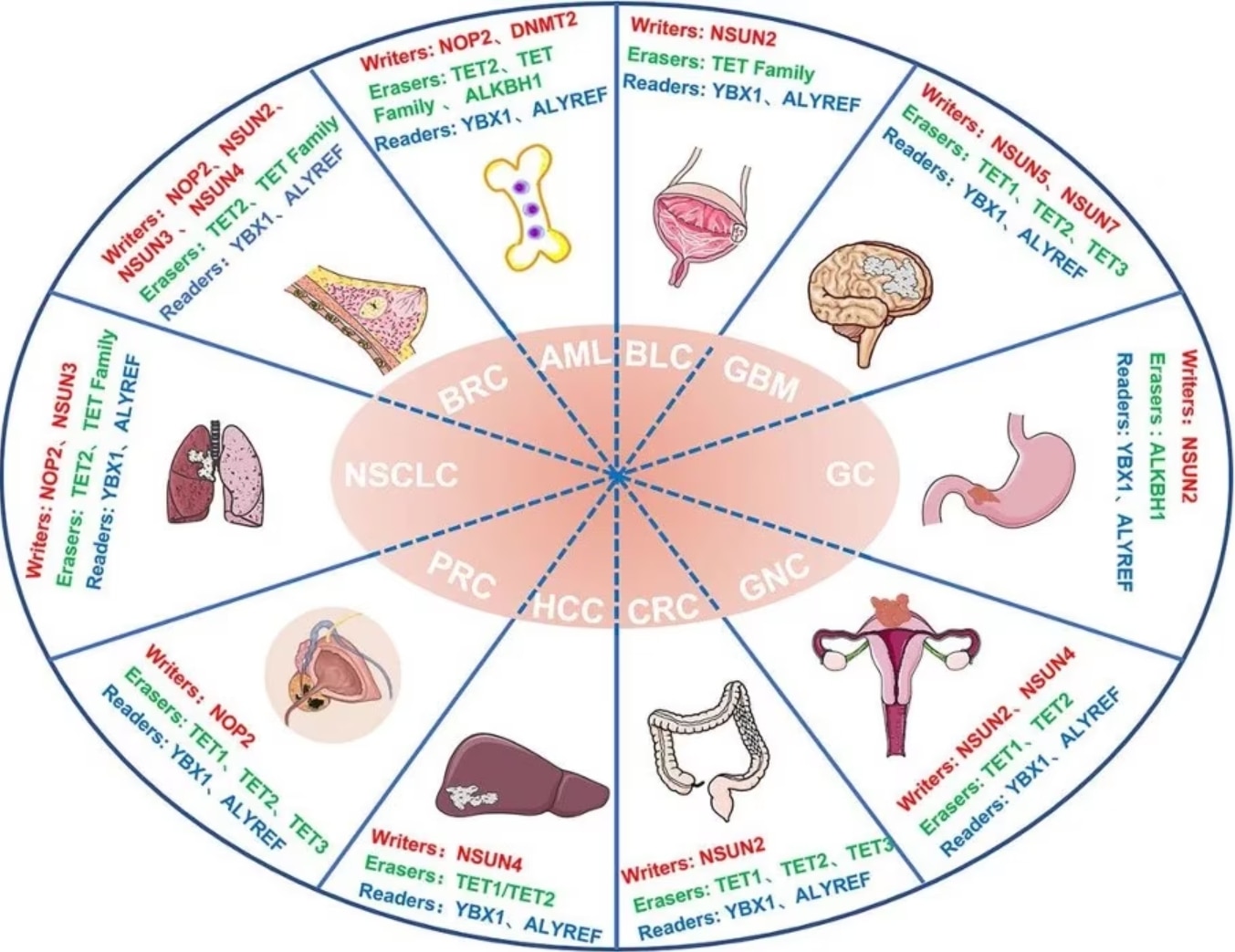
コントロールの正常組織と比較して、肝臓がん組織のmRNAは多くの部位が有意にm5Cメチル化修飾され、独特のメチル化プロファイルを示します[15]。ある研究では、その具体的なメカニズムは不明なものの、NSUN4・ALYREFの高発現と肝臓がん患者の予後不良には相関があることが認められています[16]。
Writerタンパク質であるNSUN2の発現亢進は、胃がんにも認められ、m5C修飾依存的なp57Kip2の発現抑制を介して胃がん細胞の増殖を促進すると考えられています[17]。さらに、Readerタンパク質であるYBX1は、m5C修飾されたmRNAを認識し、ELAVL1を動員してHDGF mRNAを安定化し、膀胱がん細胞の増殖や転移を促進します[18]。
その他の研究では、m5C修飾レベルが低い甲状腺乳頭癌(PTC)患者は、休止状態にあるCD4+メモリーT細胞やCD8+T細胞の組織浸潤レベルが高く、予後が良好であり、m5C修飾レベルの高いPTC患者は、活性化状態にあるNK細胞や単球の組織浸潤レベルが高く、予後が不良であることが示されています[19]。
|
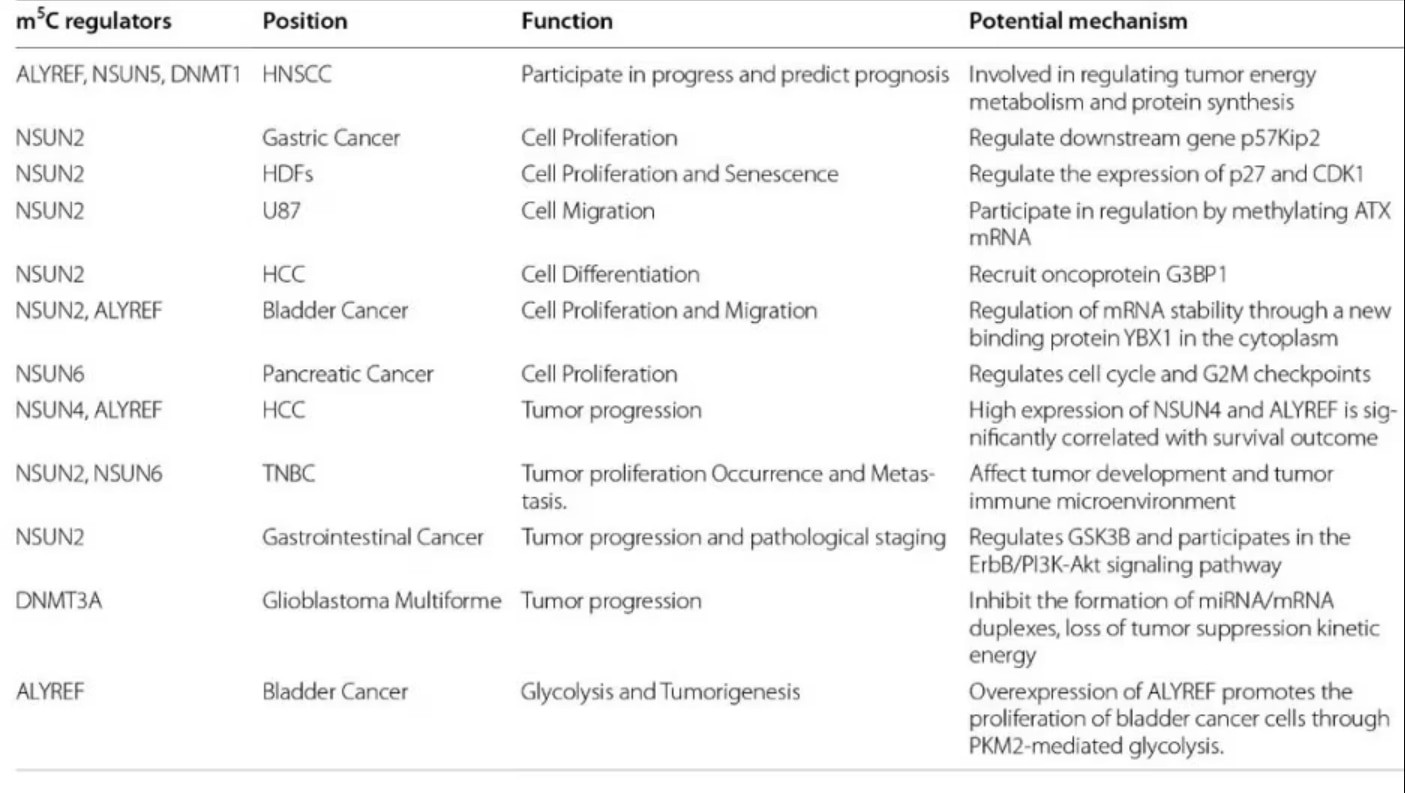
表1. m5C制御因子とがん(引用元:Song et al. 2022[2]) |
m5C修飾の研究は現在も活発に行われており、特にがん領域においては、新規治療ターゲットの同定や、診断・治療・予後予測に関する新たな知見が次々と得られています。
プロテインテックのm5C修飾研究用抗体
| 5-methylcytosineモノクローナル抗体(カタログ番号:68301-1-Ig) | NSUN2モノクローナル抗体(カタログ番号:66580-1-Ig) |
|
図4. HeLa細胞から全DNAを単離し、異なる濃度でニトロセルロースメンブレンにドット状に滴下した。メンブレンをBSAでブロッキング処理し、5-methylcytosine抗体(カタログ番号:68301-1-Ig、希釈倍率1:5000)を使用してブロッティングを実施した後、HRP標識ヤギ抗マウス二次抗体とインキュベーションし、ECL基質で発色させた。同時に、マウスIgG2bアイソタイプコントロール(カタログ番号:66360-3-Ig)を使用して同じ条件でコントロール実験を実施した。 |
図5. NSUN2抗体(カタログ番号:66580-1-Ig、希釈倍率1:40000)を使用したHepG2細胞のウェスタンブロット。NSUN2過剰発現HepG2細胞と、ネガティブコントロールとしてshRNA導入HepG2細胞を使用した。 |
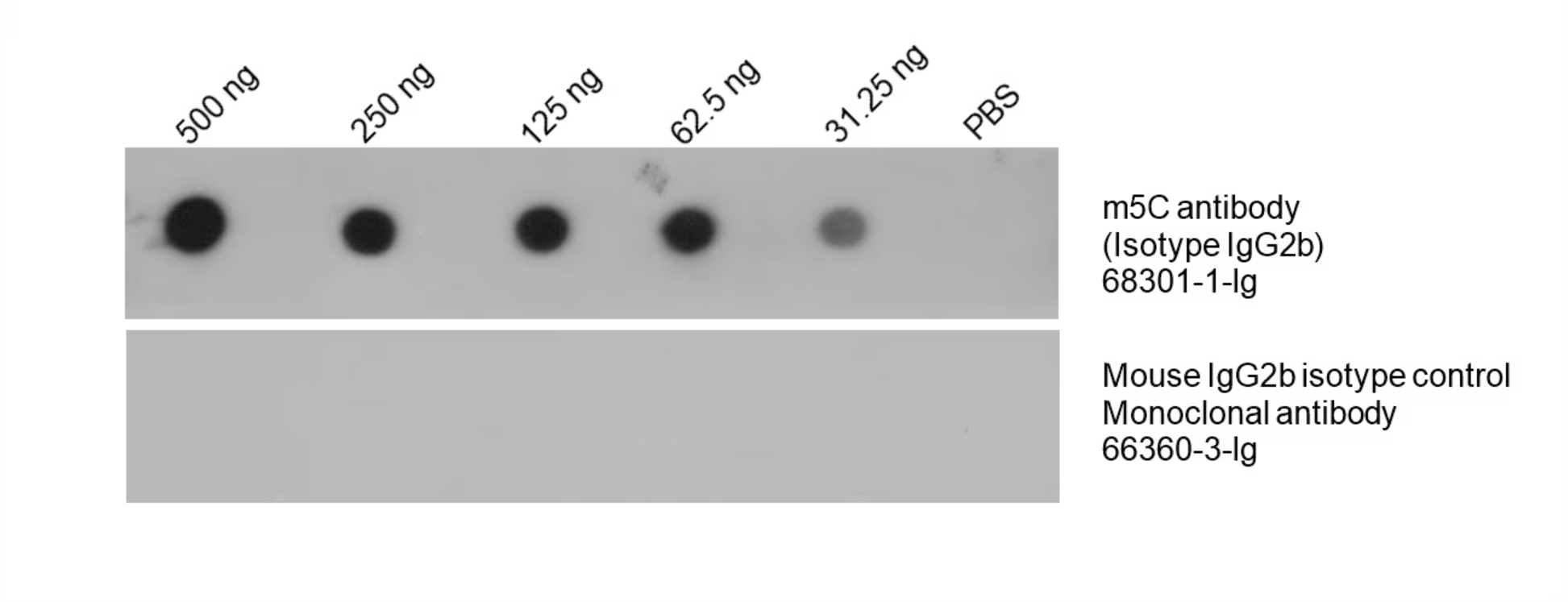
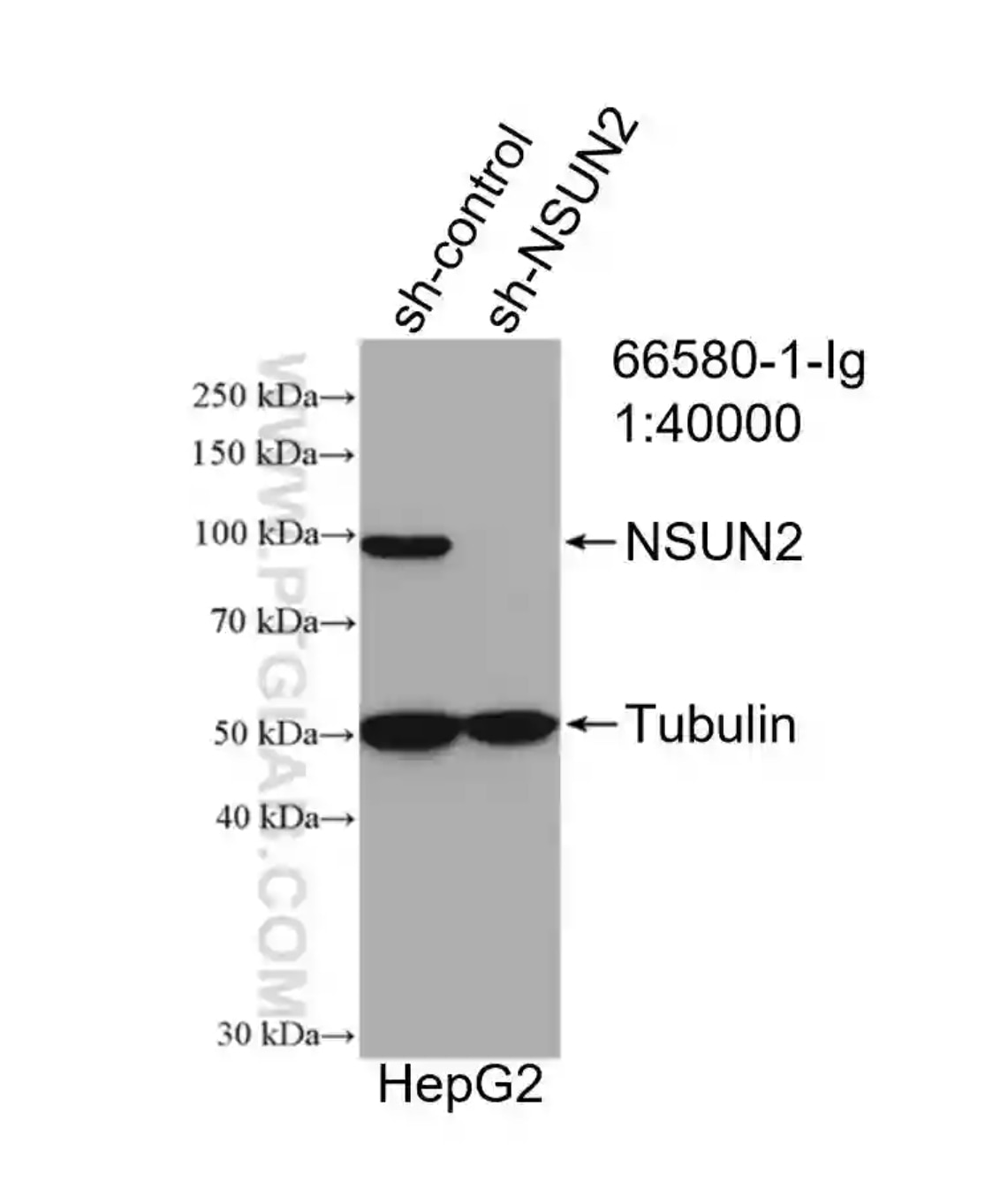
表2. 関連抗体一覧
|
ターゲット |
カタログ番号 |
Host/Type |
アプリケーション |
|
68301-1-Ig |
Mouse monoclonal |
WB, IHC, IF, RIP, Dot Blot, ELISA |
|
|
16690-1-AP |
Rabbit polyclonal |
WB, IHC, IF/ICC, RIP, ELISA |
|
|
24206-1-AP |
Rabbit polyclonal |
WB, IP, CoIP, ChIP, RIP, ELISA |
|
|
68485-1-Ig |
Mouse monoclonal |
WB, IF, ELISA |
|
|
19221-1-AP |
Rabbit polyclonal |
WB, IF, ELISA |
|
|
20954-1-AP |
Rabbit polyclonal |
WB, IHC, IF/ICC, IP, ChIP, RIP, ELISA |
|
|
26971-1-AP |
Rabbit polyclonal |
WB, IHC, IF, IP, CoIP, ChIP, ELISA |
|
|
14939-1-AP |
Rabbit polyclonal |
WB, ELISA |
|
|
60064-1-Ig |
Mouse monoclonal |
WB, IHC, IF, CoIP, ELISA |
|
|
11910-1-AP |
Rabbit polyclonal |
WB, IHC, IF/ICC, FC (Intra), IP, CoIP, ChIP, RIP, ELISA, CLIP |
|
|
20854-1-AP |
Rabbit polyclonal |
WB, IHC, IF/ICC, FC (Intra), IP, CoIP, RIP, ELISA |
|
|
66580-1-Ig |
Mouse monoclonal |
WB, IHC, IF/ICC, IP, CoIP, ELISA |
|
|
15449-1-AP |
Rabbit polyclonal |
WB, IHC, IF, RIP, ELISA |
|
|
17240-1-AP |
Rabbit polyclonal |
WB, IHC, IF, RIP, ELISA |
|
|
17546-1-AP |
Rabbit polyclonal |
WB, IHC, ELISA |
|
|
67024-1-Ig |
Mouse monoclonal |
WB, IHC, IF, ELISA |
|
|
28045-1-AP |
Rabbit polyclonal |
WB, IP, CoIP, ELISA |
|
|
21207-1-AP |
Rabbit polyclonal |
WB, IHC, IF/ICC, FC (Intra), CoIP, ChIP, RIP, ELISA |
|
|
20339-1-AP |
Rabbit polyclonal |
WB, IHC, IF/ICC, IP, CoIP, ChIP, RIP, ELISA |
References: