
ミトコンドリアのストレス応答「UPRmt」の解明
これまで解明されていなかったミトコンドリアのUPRのメカニズムを明らかにしたNature誌掲載論文を紹介します。
Joris Frenz著(ドイツ癌研究センター、博士研究員、Proteomics and Cancer Cell Signaling研究室、ドイツ/ハイデルベルク)
本稿ではF X R Sutandy, I Gößner, G Tascher, C MünchらによるNature論文「A cytosolic surveillance mechanism activates the mitochondrial UPR(PMID:37286597)」を要約して解説します。
細胞生物学という複雑な分野において、タンパク質恒常性の維持、すなわちプロテオスタシス(Proteostasis)は細胞が正常に機能を発揮し生存していくうえで極めて重要な機構です。タンパク質は細胞内において極めて多岐にわたる必要不可欠な役割を果たすため、タンパク質のフォールディング障害や機能不全が生じると細胞ストレスや細胞機能障害が発生します。ここで重要となるのは、ミトコンドリアには異常タンパク質の蓄積等のストレスによって惹起されるUPRmt(Mitochondrial unfolded protein response)と呼ばれる固有の品質管理機構が備わっているという点です。
Sutandyらによる論文「A cytosolic surveillance mechanism activates the mitochondrial UPR」では、UPRmtのシグナル伝達機構やミトコンドリアのプロテオスタシス維持における役割等を調査し、UPRmtの非常に興味深い機構を明らかにしています。
|
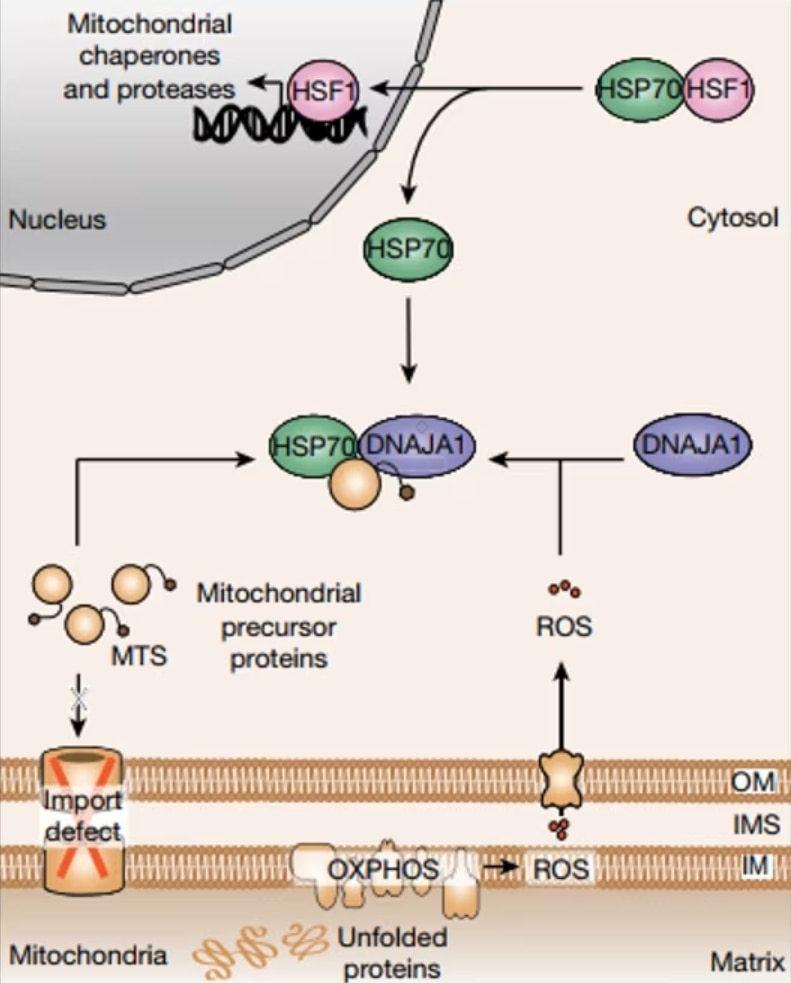
図1. 細胞質におけるUPRmt監視機構とそれに続くUPRmt関連因子転写活性化の模式図。 |
UPRmtとは?
UPRmtとは、核における特定の転写を活性化することで、タンパク質の異常蓄積に起因する毒性からミトコンドリアを保護する細胞内プロセスです。ミスフォールドタンパク質やアンフォールドタンパク質の蓄積を介して「ミトコンドリアのミスフォールディングストレス(MMS:Mitochondrial misfolding stress)」が発生すると、UPRmt機構が働いてミトコンドリアという重要なオルガネラのプロテオスタシスが適正な状態に回復します。UPRmtの重要性は広く立証されているものの、ミトコンドリアのMMSを介した核へのシグナル伝達メカニズムはいまだに全容が明らかになっていません。
ミトコンドリア活性酸素種(mtROS:Mitochondrial Reactive Oxygen Species)
研究者らが第一段階として、UPRmtを誘導しRNAseqを実施したところ、酸化ストレス応答に関与するタンパク質の転写が有意に上昇することが明らかになりました。さらに、MMSが発生した細胞においてミトコンドリア活性酸素種(mtROS)の増加が認められました。ミトコンドリアは正常に機能する過程で副生成物としてROSを産生することが判明していますが、その一方でこの研究ではmtROSの酸化ストレスがUPRmtにおいて重要な役割を果たすことが示唆されました。MMS発生時、mtROS産生が増加するとmtROSは細胞質に拡散し、ストレスシグナルを伝達するシグナル伝達分子として作用します。さらに、HSP70の補助因子として働くHSP40のファミリータンパク質DNAJA1をノックダウンすると、GTPP(ミトコンドリアHSP90阻害剤)によって誘導したMMSのストレス環境下でミトコンドリアプロテアーゼLONP1の転写活性化が抑制されることが明らかになりました。この発見はmtROSを介したUPRmtにおいて、DNAJA1が重要な役割を果たすことを示しています。
細胞質におけるミトコンドリア前駆タンパク質の蓄積(c-mtProt)
mtROSの解析と並行して、本研究では細胞質に蓄積するミトコンドリア前駆タンパク質(c-mtProt)が果たす、UPRmt活性化の極めて重要なシグナルとしての役割についても解明を試みています。ミトコンドリアタンパク質は細胞質でタンパク質前駆体として生合成され、その後ミトコンドリア内部に輸送されてプロセシングを受け、成熟型機能性タンパク質へと変換されます。酵母のMMSでは、ミトコンドリアへのタンパク質輸送不全が生じると細胞質に前駆体タンパク質が蓄積し、プロテオスタシスを回復させる必要があることをシグナルとして伝達します。類似機構はヒト細胞にも存在し、シクロヘキシミドで翻訳機構を阻害して細胞質のミトコンドリア前駆タンパク質の蓄積を防ぐと、UPRmt活性化は有意に抑制されます。同様の効果はミトコンドリア生合成を制御する転写因子であるNRF1を枯渇させた際にも認められており、UPRmtシグナル伝達はミトコンドリア前駆体タンパク質の蓄積を選択的に感知する経路であることが示されています。
ミトコンドリアと細胞質のプロテオスタシス:両者をつなぐ監視機構
mtROSとc-mtProtシグナルの両者が統合することで、UPRmtの活性化が誘導されます。ミトコンドリアから放出されたmtROSは細胞質に存在するHSP40タンパク質のDNAJA1を酸化し、c-mtProtへの細胞質HSP70の動員を促します。その結果、HSP70と結合していた転写因子HSF1はHSP70から遊離し、核へと移行してUPRmt関連遺伝子の転写を活性化します。ここで重要なポイントは、ただROSを誘導しただけではUPRmtが観察されなかったという点です。mtROSとc-mtProtが緊密に連携した細胞質監視機構は、ミトコンドリア内のプロテオスタシスの回復を担保する重要な仕組みといえます。
酵母における典型的なUPRmtの存在は報告されていませんが、酵母やその他の生物モデルにおいて、c-mtProtの蓄積が引き金となって細胞質プロテオスタシスがリモデリングされる現象が観察されています。ヒトの場合は、より複雑な細胞質監視システムが複数のシグナルを統合し、UPRmtを活性化する役割を担います。ミトコンドリアと細胞質プロテオスタシス間の連携メカニズムは、細胞質プロテオスタシスの崩壊とミトコンドリア機能低下が関係する疾患を解明する糸口を開くと考えられます。
将来的な方向性
この研究で示された発見は、細胞内プロテオスタシスを解明しその機構を制御するための様々な可能性を示唆しています。UPRmtの研究が進めば、細胞が生存するうえで必須の過程であるタンパク質フォールディングとその機能の繊細な均衡を保つ、細胞の複雑な機構が明らかになると考えられます。UPRmt活性化に関与するシグナル伝達経路の解明は、ミトコンドリアや細胞質プロテオスタシスの機能不全と関連する疾患を治療するための知見をもたらす可能性を秘めています。
さらにUPRmtと細胞質プロテオスタシスの関係の解明は、加齢や加齢性疾患に関連する細胞プロセスの解明にもつながります。ミトコンドリア機能低下とプロテオスタシス不全は加齢の重要な指標です。ミトコンドリア・細胞質のタンパク質恒常性を維持する機構の解読は健康長寿のための手法を構築する糸口を開くと考えられます。
ミトコンドリア活性酸素種(mtROS)と細胞質に蓄積するミトコンドリア前駆タンパク質のシグナル統合機構は、ミトコンドリアと細胞質プロテオスタシスの緊密なネットワークの存在を裏付けています。一連の発見は、細胞内プロテオスタシスと、身体の生理的機能や病態形成との関係性を探る研究を前進させる足掛かりとなります。
この論文に使用されたプロテインテック製品
| COX5Bポリクローナル抗体 | (カタログ番号:11418-2-AP) |
| DNAJA1ポリクローナル抗体 | (カタログ番号:11713-1-AP) |
| HSP70ポリクローナル抗体 | (カタログ番号:10995-1-AP) |
| LONP1ポリクローナル抗体 | (カタログ番号:15440-1-AP) |
|
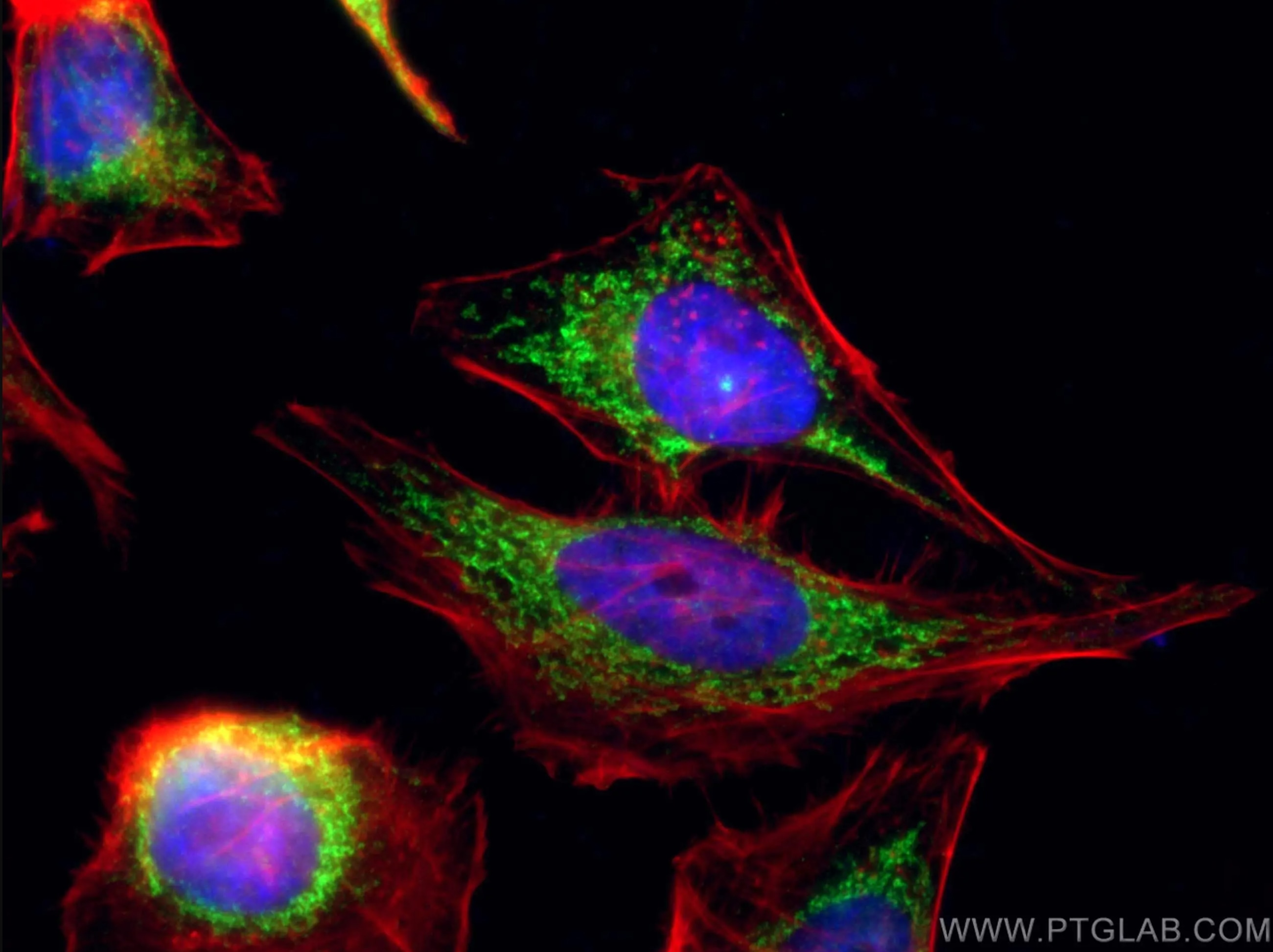
注目製品 COX5Bポリクローナル抗体(カタログ番号:11418-2-AP)
COX5B抗体(カタログ番号:11418-2-AP、希釈倍率1:100)、CoraLite® 488標識AffiniPure Goat Anti-Rabbit IgG(H+L)を使用したHeLa細胞の免疫蛍光染色像。赤:ファロイジン(Phalloidin)で染色したF-actin。細胞:4% PFA固定。 |
|
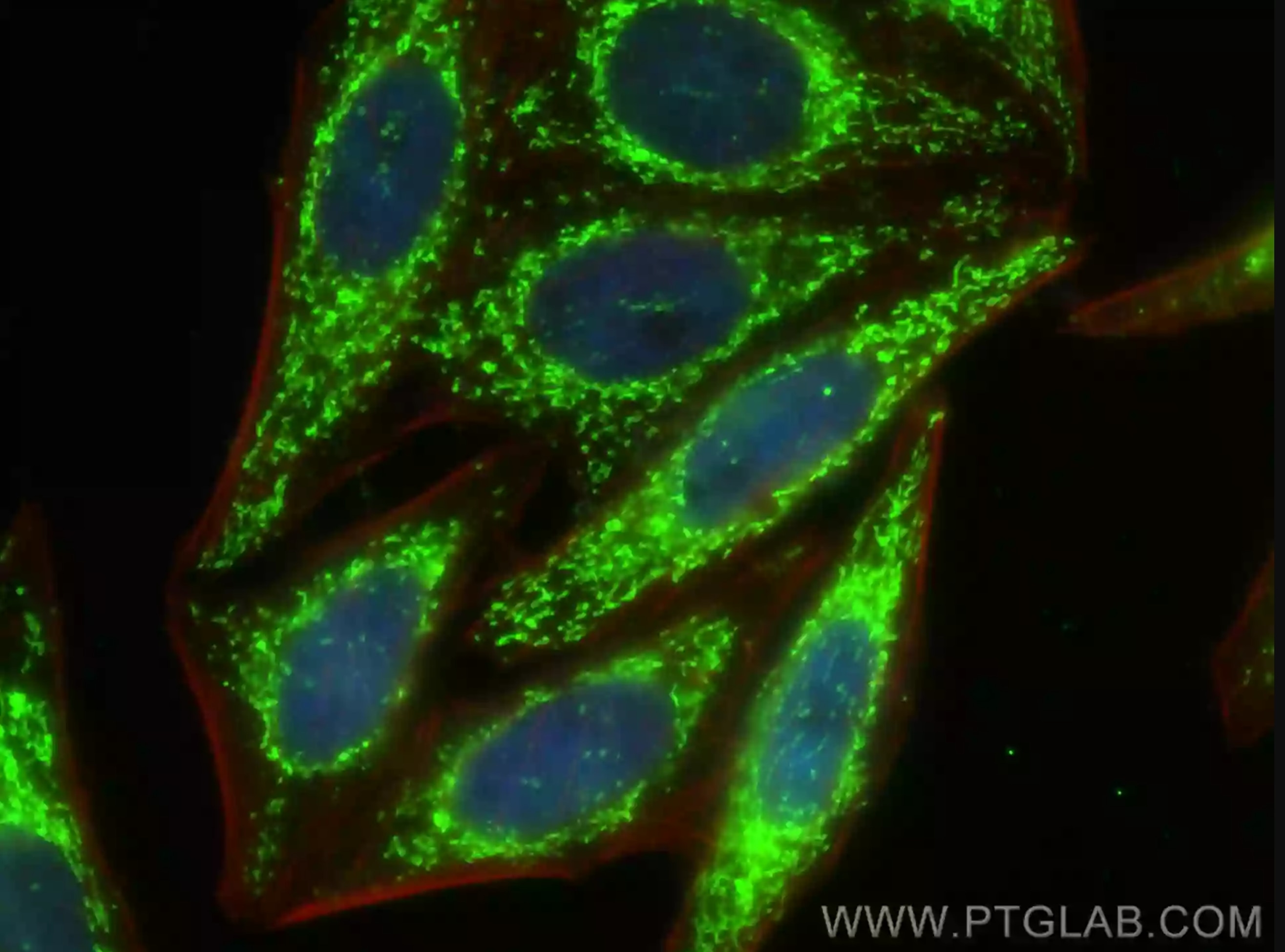
注目製品 LONP1ポリクローナル抗体(カタログ番号:15440-1-AP)
LONP1抗体(カタログ番号:15440-1-AP、希釈倍率1:200)、CoraLite® 488標識AffiniPure Goat Anti-Rabbit IgG(H+L)を使用したHepG2細胞の免疫蛍光染色像。赤:CL594-Phalloidinで染色したActin。細胞:4% PFA固定。 |
プロテインテックの「特集:ミトコンドリアマーカー」
引用数の豊富なミトコンドリア研究用抗体を中心に幅広い関連製品を紹介しています。
| BAXポリクローナル抗体 | (カタログ番号:50599-2-Ig) |
| TOM20ポリクローナル抗体 | (カタログ番号:11802-1-AP) |
| COXIVポリクローナル抗体 | (カタログ番号:11242-1-AP) |
| SOD2ポリクローナル抗体 | (カタログ番号:24127-1-AP) |