特集 : ミトコンドリアマーカー
細胞シグナリングとエネルギーバランスの中心
目次背景 |
背景
ミトコンドリアは、重要な細胞小器官 (オルガネラ) の一つであり、その役割には、細胞のエネルギーバランスの維持、細胞死プロセスの調節、細胞の酸化的ストレスの生成、カルシウム恒常性の維持等が含まれます。
近年の研究では、癌、アポトーシス、オートファジー、低酸素症を含む様々な現象への関連することが示されており、現在、「ミトコンドリア」は研究の最前線となっています。今後、タンパク質が細胞内でどの位置に局在するかを知ることによって、まだ明らかにされていないタンパク質の役割の解明につながる可能性があります。
プロテインテックは、ミトコンドリア研究に必要とされるおそらくすべての抗体を供給します。
ブログ:ミトコンドリア : 細胞シグナリングとエネルギーバランスの中心
|
ミトコンドリアマーカー
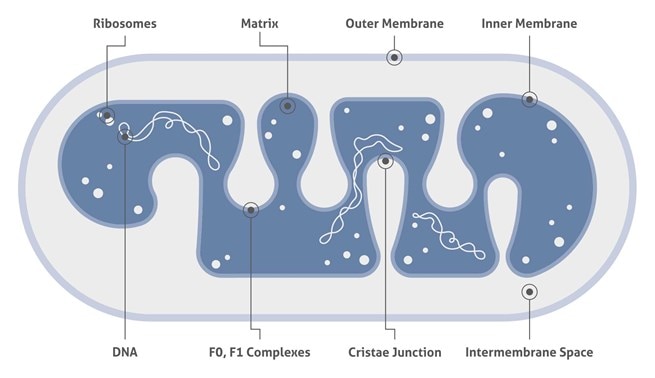
ミトコンドリアは、内外の膜、膜間隙、クリスタ、マトリックスから構成され、核とは分離した独自のDNA (ミトコンドリアDNA) を含みます。以下の表に示すオルガネラ特異的マーカー抗体を使用し、タンパク質の共局在化をみることによって、対象のミトコンドリアタンパク質の細胞内位置を確認することができます。
| HSPD1 | AIFM1 |
| ミトコンドリアマトリックスおよびミトコンドリア内膜のマーカー | ミトコンドリア膜間腔 (inter-membrane space) マーカー |
 |
 |
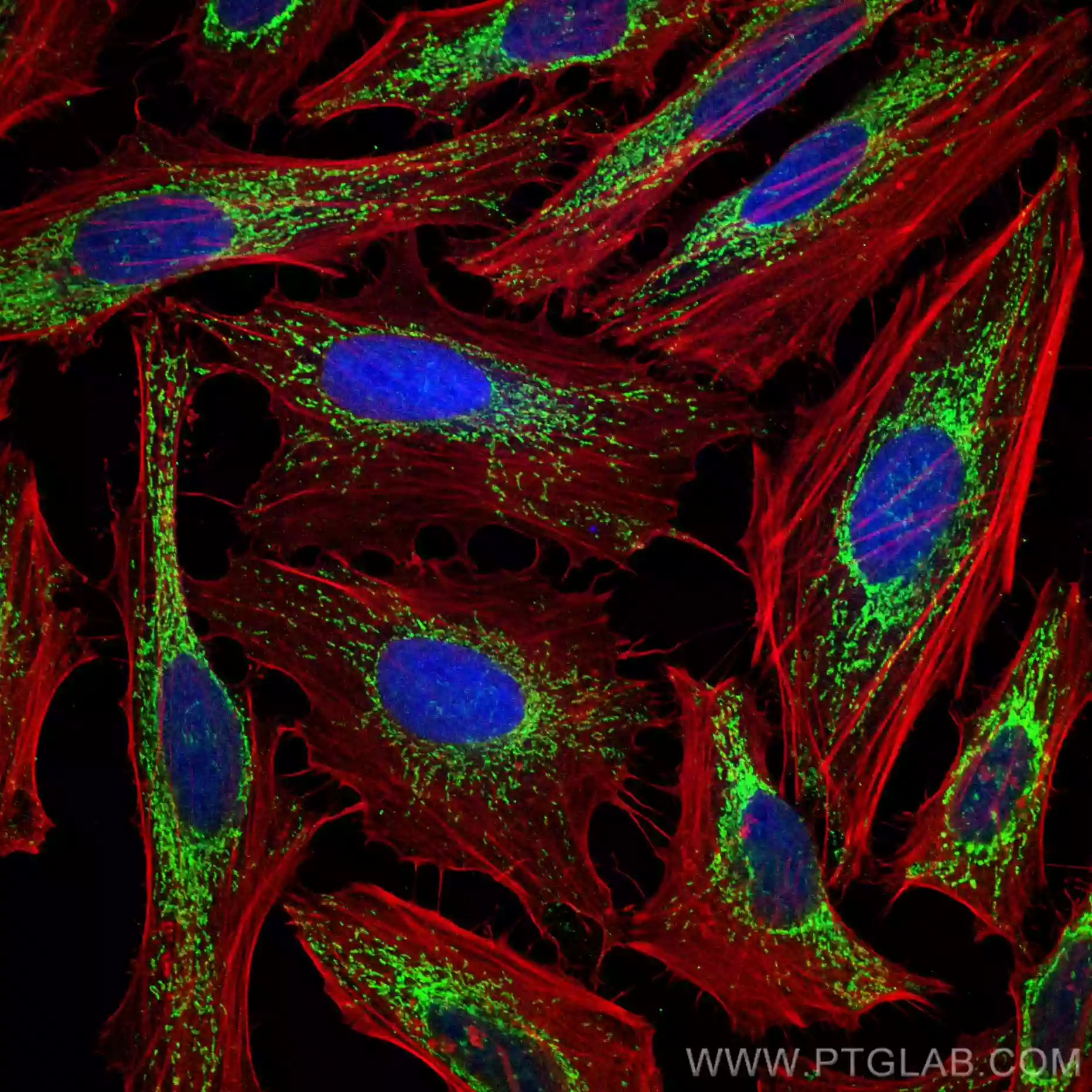
| HSPD1抗体 (品番:66041-1-Ig、希釈倍率 1:100) および CoraLite®488標識 AffiniPure Goat Anti-Mouse IgG(H+L)を用いた、 4% PFA固定Hela 細胞の免疫蛍光染色。赤色は、CL594-Phalloidin。 | AIF 抗体 (品番:17984-1-AP、希釈倍率 1:200) および Alexa Fluor 488標識ヤギ抗ウサギ IgGを用いた、4% PFA固定 HeLa 細胞の免疫蛍光染色。赤色は、CL594-Phalloidin。 |
| SCOT (OXCT1) |
TOMM40 |
| ミトコンドリアマトリックスマーカー | ミトコンドリア外膜マーカー |
 |
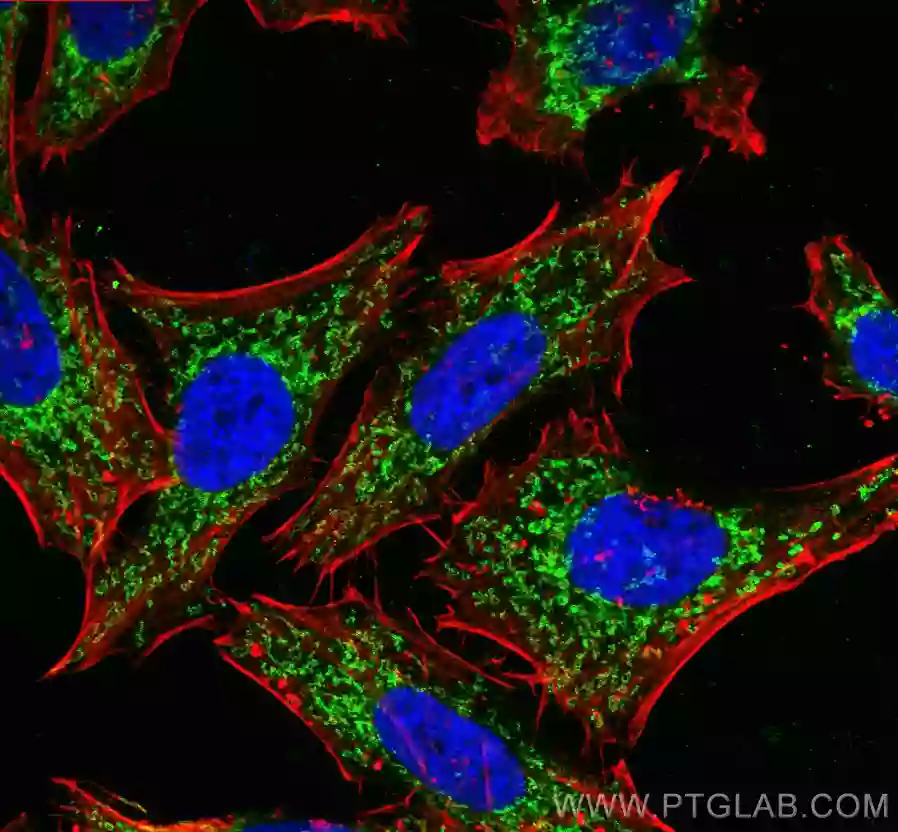 |
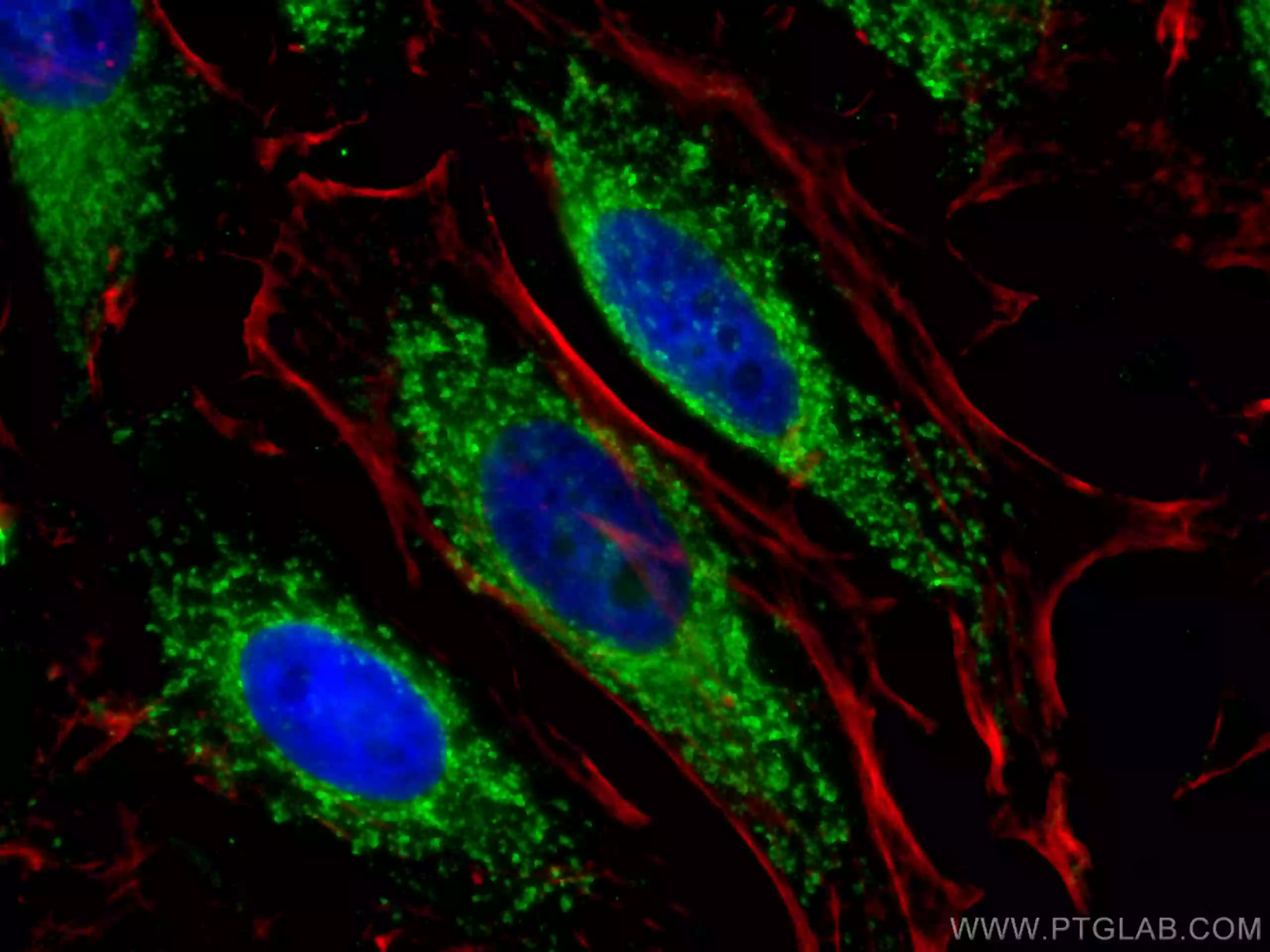
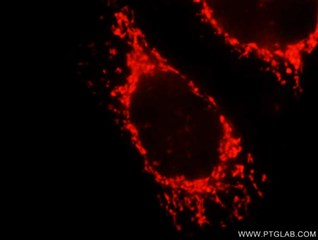
| OXCT1抗体 (品番:12175-1-AP、希釈倍率 1:25) および ローダミン標識ヤギ抗ウサギ IgG (赤色) を用いた、 MCF-7 細胞の免疫蛍光染色。 | TOMM40抗体 (品番:18409-1-AP、希釈倍率 1:400) および CoraLite®488標識ヤギ抗ウサギIgGを用いた4% PFA固定HepG2細胞の免疫蛍光染色。赤色は、CL594-Phalloidin。 |
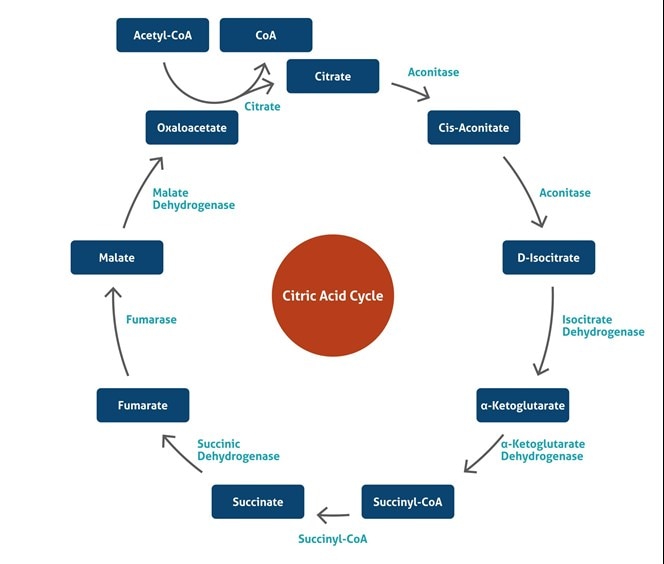
クエン酸回路 (Citric Acid Cycle)
クエン酸回路は、TCA回路またはKrebs回路としても呼ばれる、好気性生物がエネルギーを生成するための一連の化学反応のことです。この反応は、炭水化物、脂肪、タンパク質に由来するアセテートを二酸化炭素へ酸化し、アデノシン三リン酸 (ATP) の形態で化学エネルギーを生成します。また、このサイクルは、他の多くの生化学反応において使用される還元剤NADHを生成すると共に、特定のアミノ酸の前駆体を提供します。真核細胞では、クエン酸回路反応は、ミトコンドリアのマトリックス中で行われます。8種類の酵素によって、アセチルCoAの形態で存在するアセテートを、水と2分子の二酸化炭素に完全に酸化されます。
関連抗体
| 抗体名 | ||
| Aconitase 2 | IDH3A | |
| Citrate synthase | MDH1 | |
| DLD | OGDH | |
| Fumarase | OGDHL | |
| IDH2 | SdhA | |
ブログ: ゲスト寄稿 | ミトコンドリアは単なる細胞の発電所ではない
|
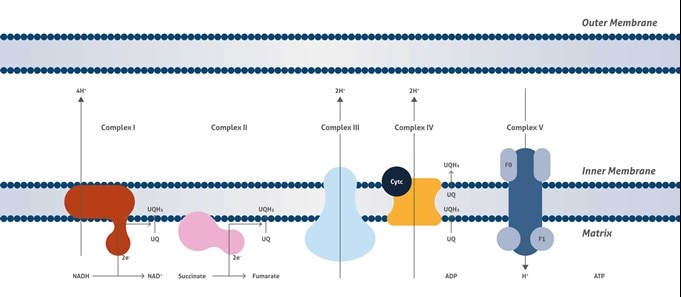
ミトコンドリア呼吸鎖複合体 (Mitochondrial Respiratory Complexes)
細胞呼吸 (Cellular respiration) は、食物からエネルギーを取り込み、生命過程にエネルギーを供給するプロセスです。ミトコンドリアの呼吸鎖は、細胞呼吸において最終的かつ最も重要な役割を果たすミトコンドリア内膜に位置するタンパク質であり、4つの大きな膜貫通タンパク質複合体からななります (複合体I、II、IIIおよびIV (CI、CII、CIII、CIV) )。それらは、CI/IIとIIIとの間のユビキノン、CIIIとIVとの間のシトクロムcを介して機能します。ミトコンドリア呼吸鎖の機能は、NADHおよびコハク酸から酸素に電子を移し、次いで内膜を横切ってプロトン勾配を作る生物学的酸化反応です。
ミトコンドリア分裂 (Mitochondrial Fission)
ミトコンドリアは、細胞機能に不可欠であり、真核生物におけるエネルギー産生、代謝産物やリン脂質、ヘムの合成、カルシウム恒常性の維持を担います。ミトコンドリアは、著しく動的なオルガネラであり、頻繁に核融合および核分裂します。核分裂と核融合という真逆のプロセスは、ミトコンドリア形態を維持し、さらに、mtDNA、代謝、生体エネルギー機能、細胞小器官数のバランスをとります。
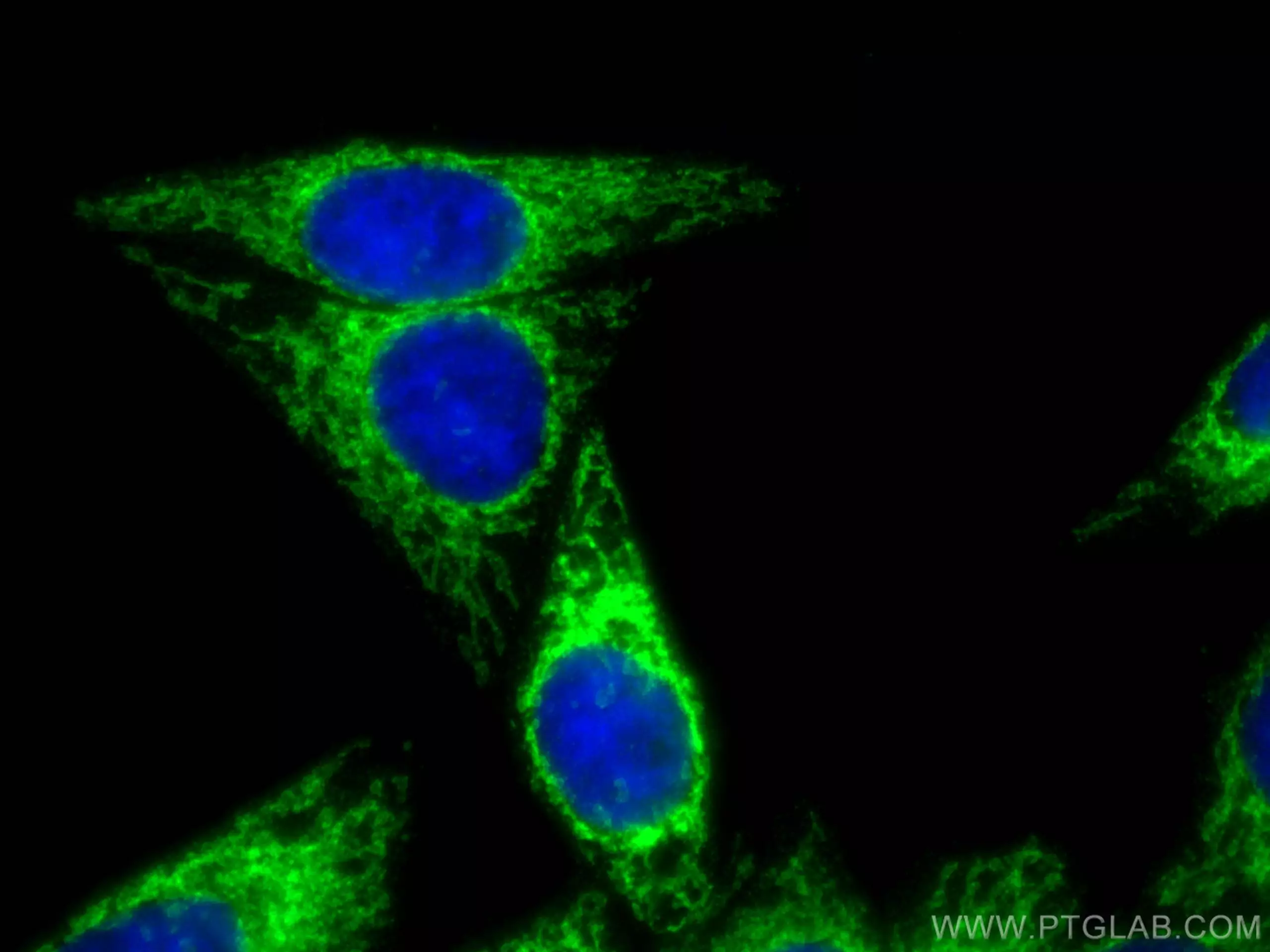
| MFF | SMCR7L |
 |
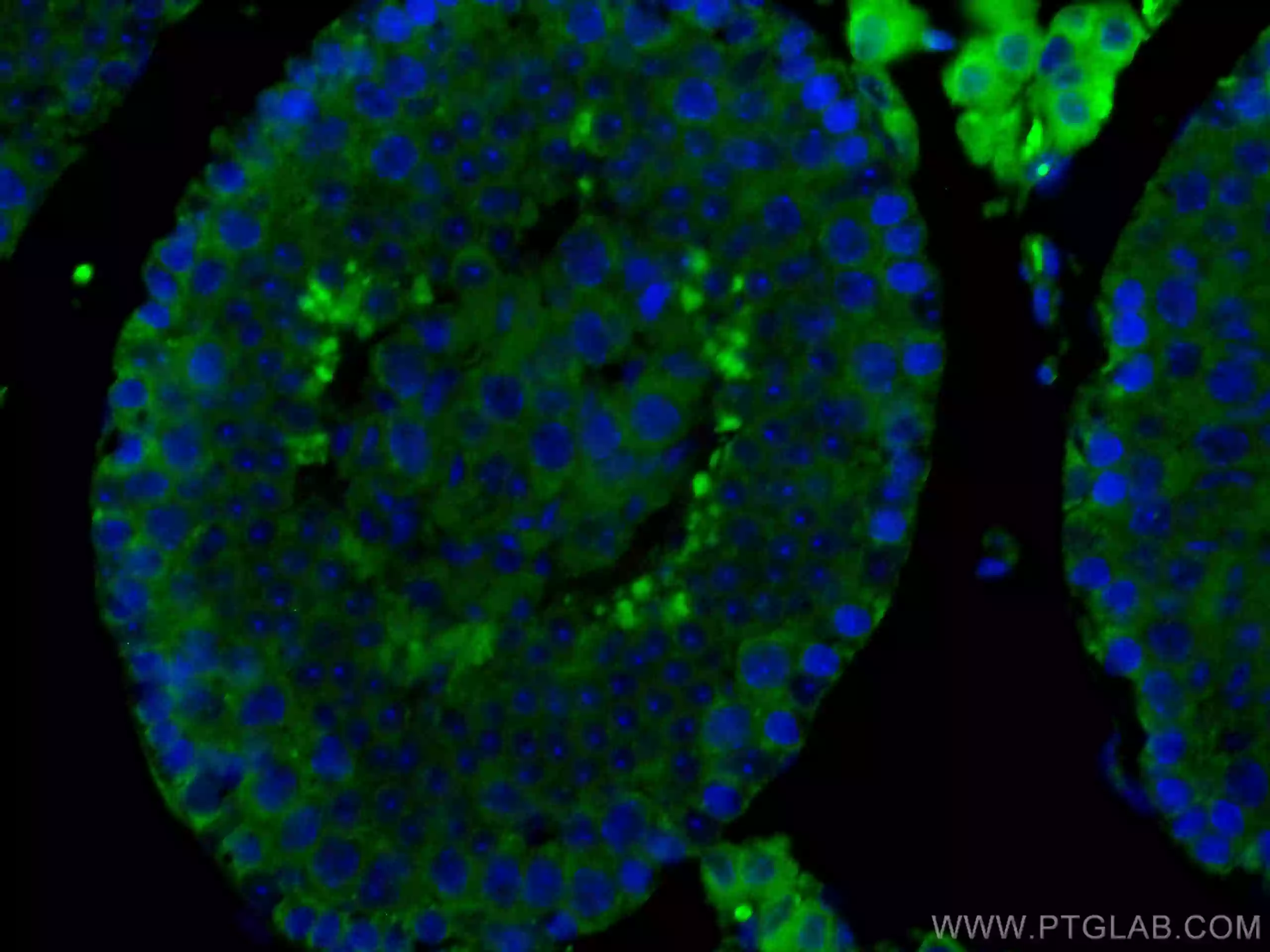 |
| MFF抗体(品番:17090-1-AP、希釈倍率 1:200) および CoraLite®488標識ヤギ抗ウサギ IgG (緑色) を用いた、 4%PFA固定 HepG2 細胞の免疫蛍光染色。 | SMCR7L抗体(品番:67808-1-Ig、希釈倍率 1:200) および CoraLite®488標識ヤギ抗ウサギIgGを用いた4% PFA固定マウス精巣組織の免疫蛍光染色。青色は、DAPI染色 (蛍光DNA色素) 。 |
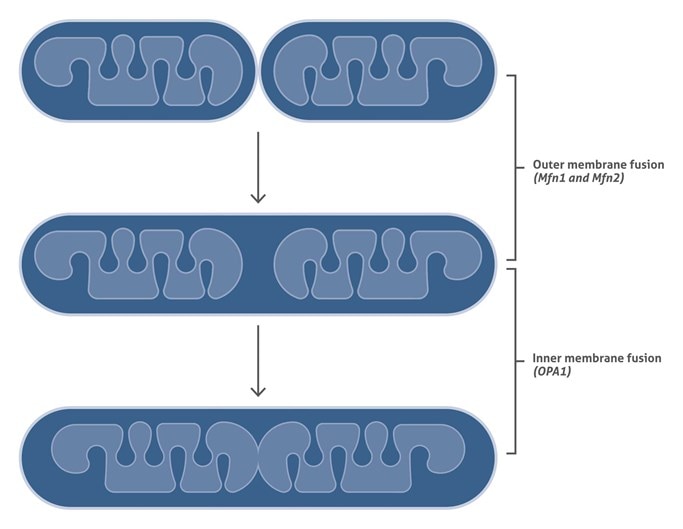
ミトコンドリア融合 (Mitochondrial Fusion)
ミトコンドリア融合は3段階で起こります:
1. ミトコンドリア外膜を介しての2つのミトコンドリアのドッキング
2. 外膜の融合
3. 内膜の融合
第1段階 (ミトコンドリア同士のドッキング) および第2段階 (外膜の融合) は、哺乳動物では Mitofusin (Mfn) タンパク質、Mfn1 および Mfn2 によって媒介されます。そして、第3段階 (内膜の融合) は、dynamin-related GTPase、OPA1によって媒介されることが知られています。
ミトコンドリア介在アポトーシス (Mitochondrial Mediated Apoptosis)
アポトーシスは、動物の発生過程および組織の恒常性維持において中心的な役割を果たす、プログラムされた細胞死 (PCD :programmed cell death) です。現在までに、2つの主要なアポトーシス経路の存在が知られており、それぞれ、ミトコンドリア (内因性経路) または細胞表面受容体 (外因性経路) によって開始されます。ミトコンドリア誘発アポトーシスは、様々な刺激に応答します。例えば、腫瘍抑制タンパク質 (例: p53) や癌遺伝子 (例: c-Myc) の活性化、DNA損傷、化学療法剤、血清飢餓や紫外線によって誘導されます。
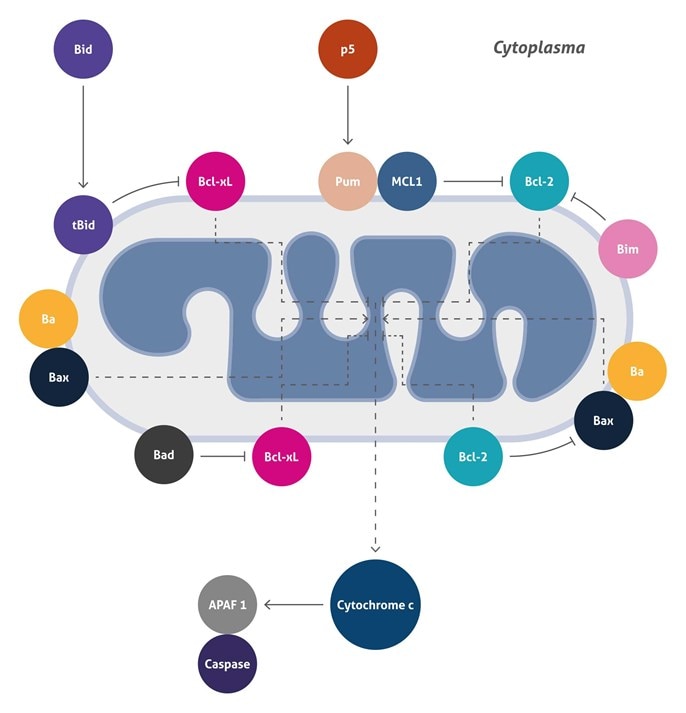
この経路は、細胞内で開始されます。ミトコンドリアの膜透過性が増加し、細胞アポトーシス分子であるチトクロームcが細胞質に放出されることから生じます。
ミトコンドリアオートファジー (Mitochondrial Autophagy)
ミトコンドリアオートファジーは、損傷を受けたミトコンドリアのオートファゴソームによる選択的除去、およびその後のリソソームによって分解するプロセス (異化作用) です。一般的に知られる機構の一つは、ミトコンドリア脱分極によって、ミトコンドリアにPINK1が安定化し、Parkinがミトコンドリアへとリクルートされることです。Parkinは、Mitofusins 1および2 (MFN1、2)、ヘキソキナーゼ (hexokinases)、TOM複合体成分、FIS1、BAK、MIROならびにVDACをユビキチン化することができ、これらは、プロテアソームを介して分解されるか、p62の結合パートナーとして機能します。p62は、オートファゴソーム膜をミトコンドリアに動員するにあたり、LC3と直接相互作用するアダプター分子として機能することが示唆されています。ParkinはAmbra1とも相互作用し、選択的マイトファジーを誘導するためにミトコンドリア周辺のPI3K複合体を活性化します。
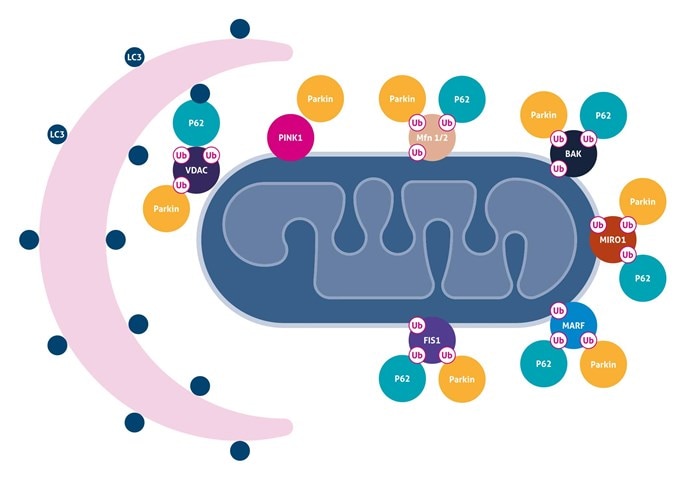
関連抗体
| 抗体名 | |
| BAK | p62 |
| Beclin 1 | Parkin |
| FIS1 | PINK1 |
| Mfn1 | VDAC1 |
| Mfn2 | VDAC2 |
| NDP52 | VDAC3 |
| OPTN |
ミトコンドリア翻訳 (Mitochondrial Translation)
ミトコンドリアは、核から分離した独自のDNA (ミトコンドリアDNA:mtDNA) を含みます。進化の過程で、ミトコンドリアがコードする遺伝子の大部分は核ゲノムに移されています。しかしながら、いくつかの遺伝子は今でもオルガネラのゲノムに保持されています。ミトコンドリアのマトリックスに収容されているミトコンドリアゲノム (mtDNA) は、ミトコンドリア内タンパク質合成に必要かつ十分であると考えられる13個のタンパク質とそのRNA分子の設計図です。ミトコンドリアタンパク質合成のために必要なその他の要素は、細胞質中で合成され、細胞質からミトコンドリアに運ばれます。ミトコンドリアタンパク質合成のプロセスの中心はミトコンドリアリボソームです。
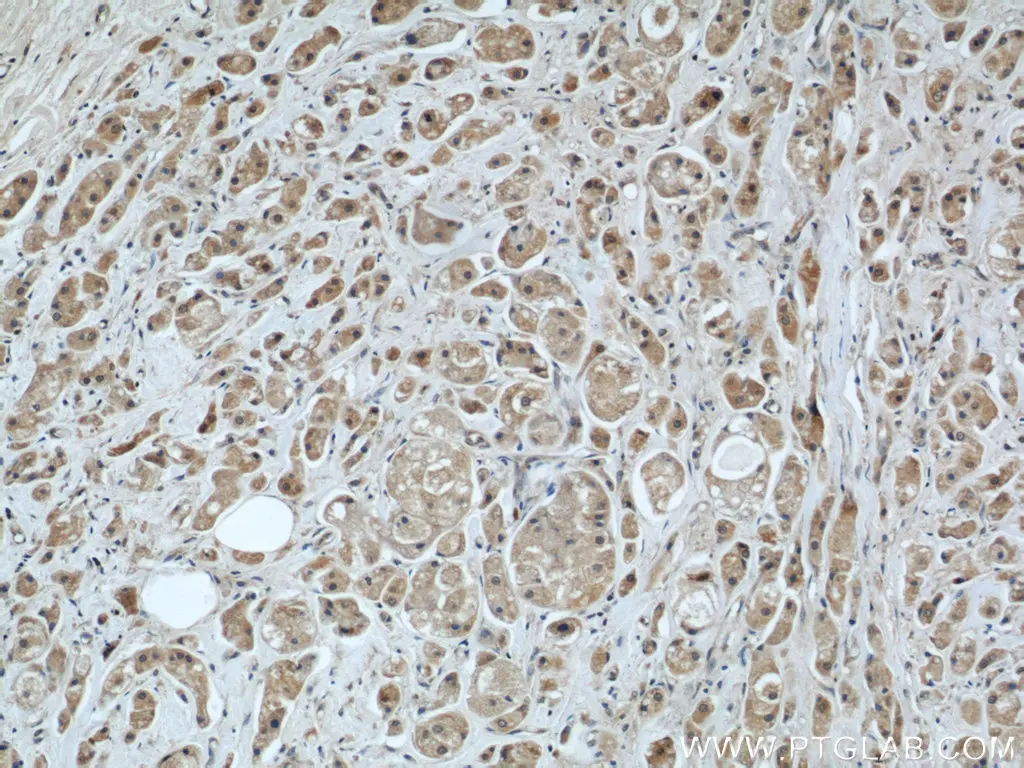
| MRPS18B | MRPS27 |
 |
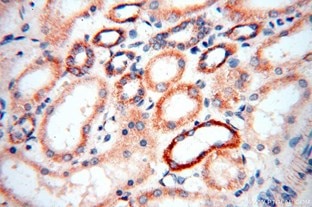 |
| MRPS18B抗体(品番:16139-1-AP、希釈倍率 1:50)を用いた、ヒト乳癌 (パラフィン包埋切片) の免疫組織化学染色 (10X) | MRPS27抗体(品番:17280-1-AP、希釈倍率 1:100)を用いた、ヒト腎臓 (パラフィン包埋切片) の免疫組織化学染色 (40X) |
ミトコンドリアはゲノム (mtDNA) とその翻訳機構を有しますが、呼吸鎖複合体等のいくつかのコアとなる構成要素を含む、ほんの少数のタンパク質のみが、mtDNA によってコードされ、オルガネラ内で合成されます。したがって、大部分のミトコンドリアを構成するタンパク質は核にコードされており、オルガネラに取り込まなければなりません。遊離リボソーム上で合成されたミトコンドリア前駆体タンパク質は、細胞質シャペロンによって誘導されて、オルガネラの表面に到達します。その後、特定のタンパク質輸送機構によってミトコンドリアに輸送され、外膜、内膜、膜間腔、またはマトリックスのどれかに蓄積されます。
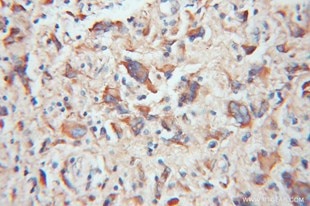
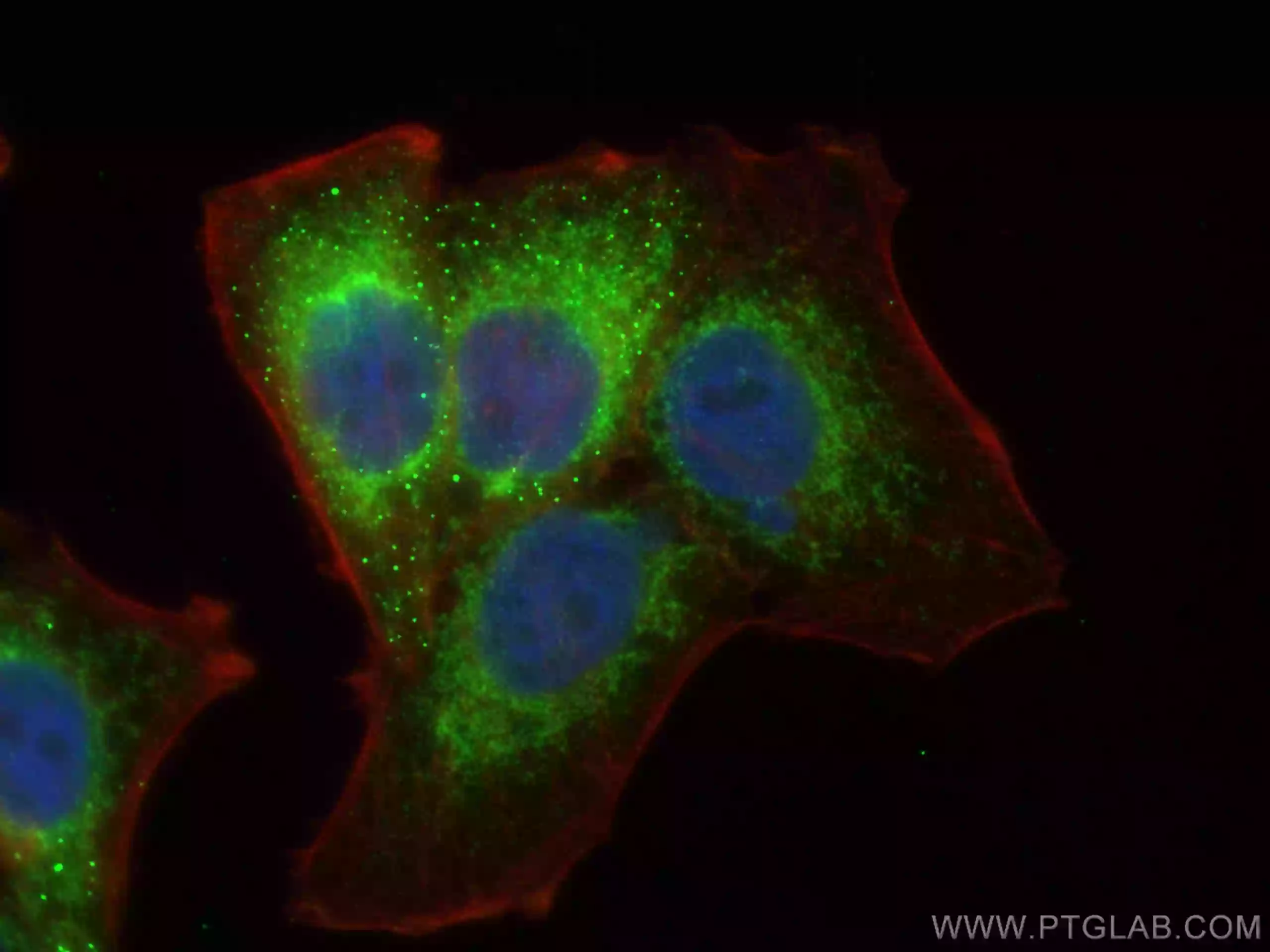
| TOM20 | TIMM23 |
 |
 |
| TOM20抗体(品番:11802-1-AP、希釈倍率 1:100)を用いた、ヒトグリオーマ (パラフィン包埋切片) の免疫組織化学染色 (10X) |
TIMM23抗体(品番:11123-1-AP、希釈倍率 1:200)およびCoraLite®488標識ヤギ抗ウサギIgGを用いた4% PFA固定HepG2細胞の免疫蛍光染色。赤色は、CL594-Phalloidin。
|
ミトコンドリアマーカーの製品フォーカスはこちら (PDF) |

